
İnsan ve insan olmayan bellek (hafıza) hayvanlar)
P. Balaban
Rusya Bilimler Akademisi Yüksek Sinirsel Aktivite ve Nörofizyoloji Enstitüsü

Bellek, gerektiğinde kullandığımız bir "depodaki" çok fazla bilgidir -?

Bellek, öğrenmenin sonucudur
Bellek, değişen koşullara uyum sağlamanın bir yoludur
Bellek: kısa süreli, orta, uzun süreli
Uzun süreli belleğin ana mekanizması sinaptik plastisitedir.

Etkinlikteki değişiklikler hakkındaki bilgiler nasıl ortaya çıkar ve sinapslarda nasıl depolanır?
Protein sentezi olmadan yeni bir hafızanın oluşması imkansızdır.
Bir protein molekülünün ortalama ömrü - gün
Hafıza, proteinlerin katılımıyla oluşuyorsa ve yıllarca saklanıyorsa, o zaman protein konsantrasyonundaki değişiklikleri sabitlemek için bir mekanizma olmalıdır.
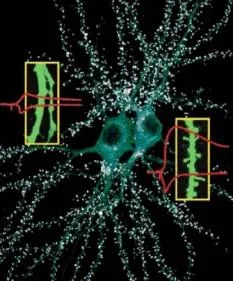
SİNAPTİK MEKANİZMALAR EVRİMSEL MUHAFAZACIDIR!
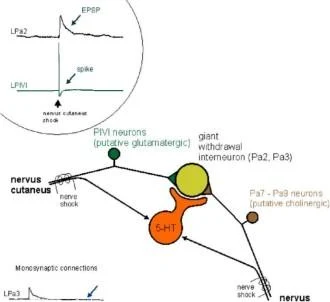
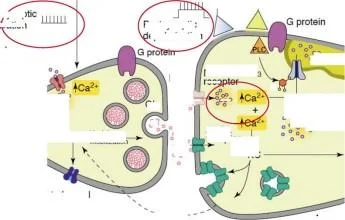
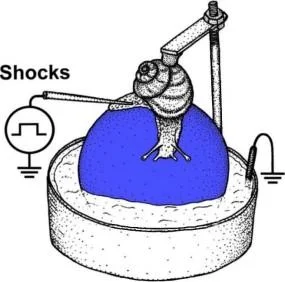
Sifon
musluk
CS

Kuyruk şoku

I Postsinaptik \lepolarizasyon
G proteini
1) Geç faz presinaptik fasihtasyon
2) Geç faz hipereksitabilite
\Aezh_0
RCA -mo S Eauon
AMPA
oa o
K* kanalı
duyusal nöronlar
Retrograd sinyal
PKC
PLTP
2. AMPA reseptör fonksiyonunun düzenlenmesi
3. Geriye dönük sinyalin uyarılması
Diğer postsinaptik değişiklikler
motor nöronlar
Roberts ve Glanzman, 2003

Hafızayı geliştiren haplar var mı?
Mart 2009'da New York Times, Dr. liderliğindeki Brooklyn Tıp Merkezi'ndeki bilim adamlarının hayatı onun için kolaylaştırdığını ciddi bir şekilde duyurdu.
Enzim protein kinaz M-zeta , uzun süreli hafıza mekanizmasının temel unsurlarından biri olarak kabul edilir (bu birkaç yıl önce kurulmuştur), ancak yazarlara göre en önemlisi, yalnızca karmaşık anıları kurtardığı için ilginçtir. , taahhüt edilen eylemler ve deneyimler hakkında ayrıntılı bilgiler şoklar. Sonuç olarak, M-zeta protein kinaz moleküllerinin seçici yıkımı ile kişi, istenmeyen olay ve deneyimleri “unutabilir” ve beyninin işleyişi bozulmaz.
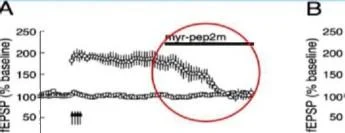
O' = S! ben S S ben
-1 0123456
Zaman (saat) Zaman (saat)
AT
10#

kızarmış ekmek
97. Gün

Tiksinme İndeksi
80
60
40
STA
gün t

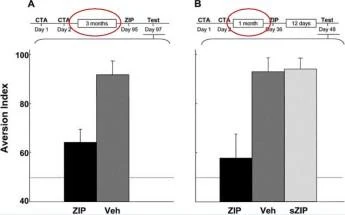
Posta Kodu
ZIP Veh sZIP
ZIP'in insular kortekste (A) çok uzun süreli CTA hafızasına etkisi ZIP/araç eğitimden 3 ay sonra uygulandı. ve hafıza 2 gün sonra test edildi. Kesikli çizgi, CS ve su için eşit tercihi gösterir, yani Al = 50. (B) ZIP/araç/karma ZIP, eğitimden 1 ay sonra uygulandı. ve hafıza 12 gün sonra test edildi. Sakarin, hem A hem de B'de CS idi.
PKmÇ
Protein kinaz M zeta
Protein kinaz C'nin (PKC) yapısal olarak aktif izoformu
Hafıza depolamaya dahil
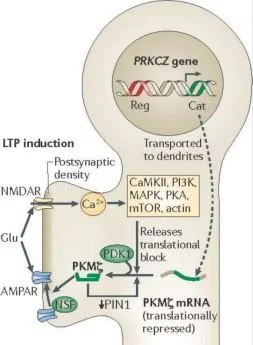
PRKCZ geni
LTP indüksiyonu
r Fbstsynaptic'
yoğunluk
i
NMDAR,

bilgi yok
Çeviri bloğunu serbest bırakır
CaMKII. PI3K, MARK. RCA, NITOR. aktin
AMPAR
PCM
dendritlere taşınır
PKMmRNA
(çeviri olarak
bastırılmış)
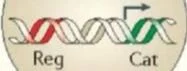
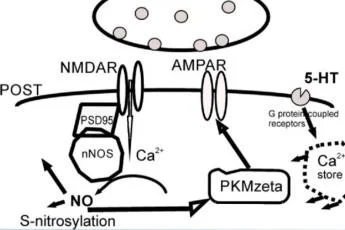
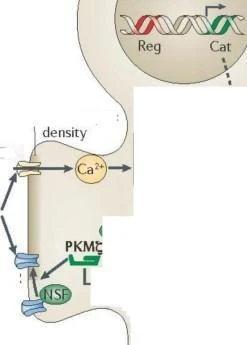
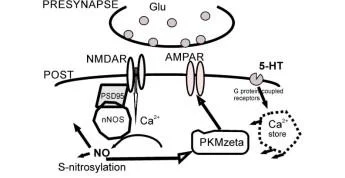
LTP'de yer almak için PKM {. Protein kinaz C, zeta (PRKCZ) genin iki promotörü vardır. biri nöronlarda bir düzenleyici alanı (Reg, kırmızı ile gösterilmiştir) ve bir katalitik alanı (Cat; yeşil ile gösterilmiştir) kodlayan ekzonlardan tam uzunlukta bir protein kinaz (PKCÇ) üretir, bir dahili promotör bir protein kinaz MÇ (PKMÇ) mRNA üretir. düzenleyici bir alan olmaksızın bir < katalitik alanı kodlayan. PKM< mRNA dendritlere taşınır ve translasyonel olarak PIN1 (NIMA1 ile etkileşen protein) tarafından baskılanır. Uzun süreli güçlenme indüksiyonu sırasında, Önce sentezlenen, PKM< bindsto ve fosfoinositite bağımlı protein kinaz 1 (PDK1) tarafından fosforile edilen ve PKMÇ PKM'nin yapısal kinaz aktivitesini artıran translasyon bloğunu serbest bırakmak için NMDAR aktivasyonu tarafından uyarılan çoklu sinyal yolakları gereklidir. < daha sonra, PKM< mesajının artan dendritik çevirisini sürdürmek için PIN1 inhibisyonu yoluyla bir pozitif geri besleme döngüsü başlatır. PKMÇ, trafik proteini N-etilmaleimide-duyarlı faktör (NsF) etkisiyle postsinaptik yoğunluktaki reseptör sayısını artırarak AMPAR yanıtlarını güçlendirir.
CaMKII, Ca2+/kalmodulin bağımlı protein kinaz II, glu, glutamat; MAPK, mitojenle aktive olan protein kinaz; mToR, rapamisinin mammahan targei'si; PI3K, fosfatidilinositol 3- kinaz, PKA, protein kinaz A
Çalışmalar, hafızanın birkaç saat - gün içinde oluştuğunu, ancak bir konsolidasyon döneminden sonra, protein moleküllerinin yerel olarak kendi kendine yeniden üretilmesiyle kalıcı olarak depolandığını göstermiştir.
Başka bir deyişle, PKMz molekülü her zaman hafızanın kritik bir bileşeni olarak kalır ve hafıza stabilitesinin temelidir.
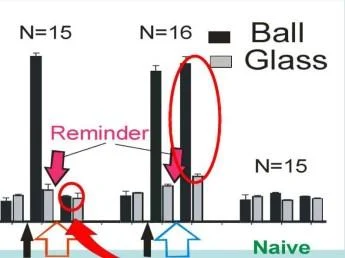
'İki farklı bağlam: 6a([andglass

BAĞLAM 1 ( TOP ÜZERİNDE )

BAĞLAM 2 (CAM ÜZERİNDE)
Hafıza depoları farklı hayvanlarda aynı mıdır?
Reninder
. deneme günleri

COMTXT 2 (CAM ÜZERİNDE1)
öğrenmeden önce
Balaban ve ark .. Frontiera in Celi Neurosci 2015
T1. a Onun öğrenmesi
T2. enjeksiyonlardan sonraki gün

TL PT Gpp
EP9P

Olmayanın anısını canlı bir organizmanın beyninde silmek veya yapay olarak yaratmak mümkün müdür?
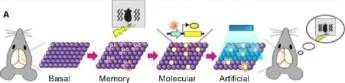
Düzey Biçimlendirme Etiketleme Aktivasyonu
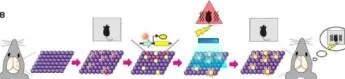
Bazal Bağlam A Moleküler Bağlam B Bağlam A Düzeyi Nötr bellek Etiketleme Işık + Şok Fasle feat bellek
Hafıza Engram Hücreleri Olgunlaştı. Neuron 67, 2 Eylül 2015 S usu mu To nega wa, Xu Lıu, S Ramırez, R Redondo
Şekil 2 Hafıza Engram Hücre Popülasyonlarının Optogenetik Mampülasyonları (A) Hafıza engram hücre popülasyonunun ışıkla aktivasyonu, hafızanın geri çağrılmasına neden oldu Bağlamsal bir korku hafızasının oluşumunda aktif olan nöronlar ChR2 ile etiketlendi. Bu nöronlar farklı bir bağlamda ışık uyarımı ile yapay olarak harekete geçirildiğinde, hayvanlar donma davranışı sergilediler, bu da korkuyla ilişkilendirilen önceki bağlamın hatırlandığını gösteriyor.
(B) Yanlış bir bağlamsal korku hafızasının oluşturulması. Nötr bir bağlamda aktif olan nöronlar, ChR2 ile etiketlendi ve daha sonra farklı bir bağlamda ışıkla yeniden aktive edildi; bu sırada memeliler, hayvanlar orijinal nötr bağlama geri döndüklerinde eş zamanlı olarak ayak şokunu düzeltti. nötr bağlamı ve aptalca şoku ilişkilendiren yanlış bir anıdan hatırlamayı gösteren korku tepkisi sergilediler.
∙ BİLİNEN:
∙ NO, sinaptik plastisite için gereklidir
∙ NO, düşük konsantrasyonlarda protein sentezini aktive ederken, yüksek konsantrasyonlarda proteinleri nitrozlayarak konformasyonlarını değiştirir.
SORU:
? HAYIR hafıza silmeye dahil mi?
OITVIOL
protein nitrozilasyonu
hÜAOÂft
• MARK, nükleer Cascade franscripHon
♦cGMP
Batı güneş$
Nitrosilasyon, bir nitrozil grubunun bir proteine translasyon sonrası eklendiği bir protein modifikasyonudur.
Joseph Loscalzo tarafından keşfedilen S-nitrosilasyon, nitrik oksidin önemli bir biyolojik reaksiyonudur; proteinlerdeki sistein kalıntıları da dahil olmak üzere tiyol gruplarının S-nitrozotiollere (RSNO'lar) dönüştürülmesini ifade eder. S-Nitrosilasyon, ana protein sınıflarının çoğunun veya tümünün dinamik, translasyon sonrası düzenlenmesi için bir mekanizmadır.
Bellek konsolidasyonu, ilk belleğin oluşumundan hemen sonra gözlemlenen varsayımsal bir süreçtir ve belleğin oluşumunu ve stabilizasyonunu yansıtır.
R -konsolidasyon, hafıza etkinleştirildiğinde gözlenen hafızanın yeniden dengelenme sürecidir ve birleştirilmiş hafızada bir değişiklik olasılığını yansıtır.
Konsolidasyon

STM
kısa süreli hafıza
Aktif durum™
OLARAK
yeniden konsolidasyon
LTM
uzun süreli hafıza
aktif olmayan durum
DIR-DİR

PROTEİN SENTEZİNE BAĞLIDIR!
PROTEİN SENTEZİNE BAĞLI DEĞİLDİR!
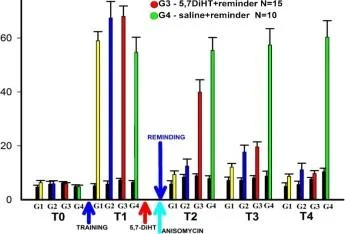
<Şek. 4 (daha küçük ( < Ejçperiment 2)
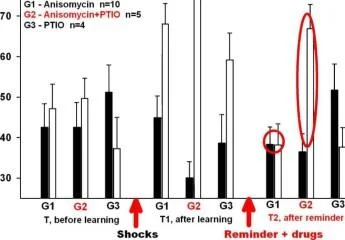
Groupl (G1) Şoklar
huni IQ
T1 T2
hatırlatıcı n
aniamisin
Grup2 (G2) Şokları
9G
T1 T2
Raminder n
nizomisin
* L-NNA
Grup3 (G3) Şoklar
mi ii
T1 T2
Hatırlatıcı
* deney günleri
L-NNA
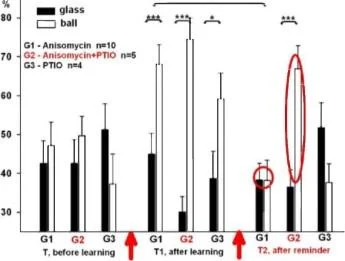
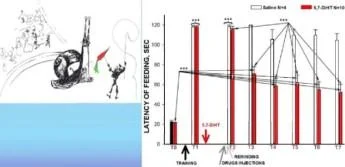
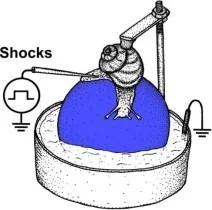
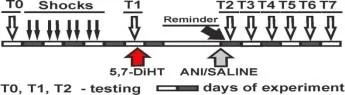
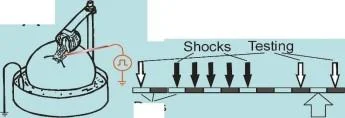
Hatırlamadan 20 dakika önce anizomisin/L-NNA enjeksiyonları ile bir bağlam şartlandırma deneyinin protokolü. T. T1, T2 - bağlam koşullandırma testleri.
<Şek.5
^minder (fcjçperiment 2)

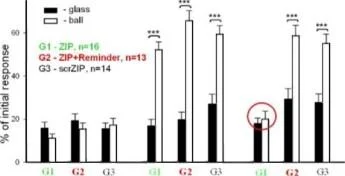
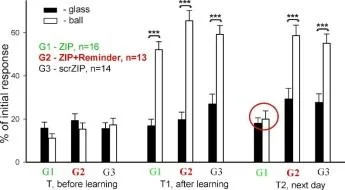
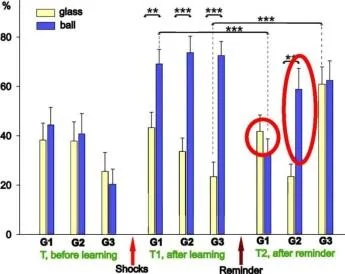
İki farklı bağlamda ölçülen üç salyangoz grubundaki geri çekilme tepkilerinin ortalama genlikleri (±SEM): bali (güçlendirilmiş bağlam) ve cam üzerinde. Grup(G1). n=8; Grup2 (G2), n=7; Grup3(G3), n=5.
Y ekseni - testten önceki uzunluğun %'si olarak dokunaç geri çekilme genliği
Şok Hatırlatma *ilaçlar
Balaban ve diğerleri Euroo. J. Nörobilim. 2014
Bellek stabilitesinin temeli, yerel konsantrasyon kendi kendine bakım özelliğine sahip PKMz molekülü ve onun homologları olabilir.
Bellek plastisitesinin temeli, bellek kararlılığı proteinlerinin işlevlerini YEREL OLARAK değiştiren nitrik oksitin yerel üretimi olabilir.
NEDEN UNUTUYORUZ?


YENİDEN KONSOLİDASYON:
YENİ PROTEİN SENTEZİNE İHTİYAÇ VAR
BELLEĞİN GÜCÜNE BAĞLIDIR
PARAMETRELERE BAĞLIDIR
HATIRLATICILAR
SÜREYE GÖRE DEĞİŞİR
BELLEK TASARRUFU
?GÜÇLENDİRME SİSTEMİNİN AKTİVİTESİNE BAĞLILIK?

BAĞLAM 1
BAĞLAM 2
80
-cam topu
EĞİTİM 5,7-DIHT
Balaban ve ark. : Bilim Raporlar, 2016
SİNYAL HAFIZASI I

BESLEME SİMÜLASYONLARI
BAĞLAM UYARMA LARI
DUYUSAL NÖRONLAR
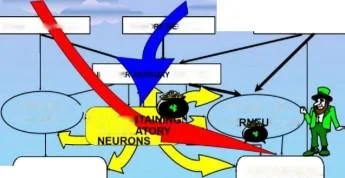
SENSO
ÜRONLAR
SİPARİŞ VERİYORUM
RONLAR
PARA ÇEKME
ARA NÖRONLARI BESLEME
5-HT-C3J
MODÜL
DUYUSAL F^KONS
BESLEME
MOTOR
MOTOR
PROGRAM
PROGRAM
°RESİNAPS
Glu
Balaban ve diğerleri, Sc. Raporlar, 2016
sonuçlar
İlk kez, hafızayı koruma sürecini başlatmak için durumsal hafızanın yeniden aktivasyonu sırasında nöral takviye sistemini aktive etme ihtiyacı gösterilmektedir.
Anahtar bir uyaran (yiyecek) için hafıza ağının nöronları, depolanması takviye sistemine bağlı olmayan ilişkisel değişikliklere uğrar.
Takviye sistemindeki nöronal aktivitenin blokajı, (1) yeniden etkinleştirme üzerine durumsal hafızanın hızlı bir şekilde yok olmasına ve (2) durumsal hafıza bileşeninin ortadan kalkması nedeniyle bir gıda uyarıcısı için hafızanın zayıflamasına yol açar.
Beynin güçlendirici sistemlerinin katılımı olmadan hafızayı tekrar tekrar etkinleştirdiğimizde unuturuz (hafızayı değiştiririz).
YÜKSEK SİNİR AKTİVİTE DERGİSİ, 2017, cilt 67, sayı 2, s. 131-140
YORUMLAR,
TEORİK MAKALELER
UDC 612.822
BELLEK MODİFİKASYONUNUN MOLEKÜLER MEKANİZMALARI
© 2017 P. M. Balaban
Federal Bütçe Bilim Kurumu Yüksek Sinirsel Aktivite ve Nörofizyoloji Enstitüsü, Rusya Bilimler Akademisi, Moskova e-posta: pmbalaban@gmail.com
28 Kasım 2016'da alındı
Yayına kabul edildi 5 Aralık 2016
Bu derlemede, kendi ve literatür verilerimize dayanarak, yeniden etkinleştirme sırasında davranıştaki uzun vadeli değişikliklerin ve bellek modifikasyonunun altında yatan, sinaptik etkinliğin düzenlenmesinin moleküler mekanizmaları hakkında bir hipotez öneriyoruz. Hipotez, atipik protein kinaz musette molekülünün, glutamat AMPA reseptörlerinin iletiminin kontrolü yoluyla sinaps etkinliğindeki uzun vadeli değişikliklerdeki rolüne ve bu moleküllerin sinapslarda üretilen nitrik oksit tarafından nitrozilasyonu olasılığına ilişkin verilere dayanmaktadır. bir sinir hücresinin aktivasyonu.
Anahtar kelimeler: nöronlar, omurgasızlar, öğrenme, Mzeta protein kinaz.
DOI: 10.7868/S0044467717020046
GİRİİŞ
Uzun süreli hafıza, beyin fizyolojisindeki en gizemli olaylardan biridir. Bugüne kadar, bu alandaki araştırmacılar, yalnızca hafızanın ve sinir sisteminin her türlü esnekliğinin yeni koşullara uyum sağlamanın temeli olduğu konusunda hemfikirdir. Uzun süreli davranış değişikliğine neden olan değişikliklerin sinir hücresinde nerede meydana geldiği ve nelerden oluştuğu konusunda tam bir fikir birliği yoktur. Bazı hafıza biçimlerinin, belirli davranışlar için omurgasızların sinir ağının anahtar nöronlarındaki uyarılabilirlikteki uzun vadeli değişiklikler düzeyinde gerçekleştirildiğine dair ikna edici kanıtlar vardır (sinaptik olmayan plastisite biçimleri [Nikitin ve diğerleri, 2013]), ancak, omurgalı hayvanlar üzerinde yapılan çalışmalarda, sinaptik temaslarda plastisite hipotezi, tüm sinir ağının tepkisinin ve davranışının modifikasyon mekanizmalarını yansıtır. Sinaps verimliliğinin kısa vadeli (onlarca dakika) düzenlenmesi için bir mekanizma olarak presinaptik plastisite kavramına dayanır [Roberts & Glanzman, 2003], uzun vadeli değişiklikler paralel olarak başlatılır ve ana süreçler uzun indüksiyonun indüklenmesidir. -term plastisite, postsinaptik nöron bölmelerinde meydana gelir [Cai ve diğerleri, 2008]. Uzun süreli bellek süreçlerine özgü moleküler sistemler için adaylar da ortaya çıkmıştır [Sacktor, 2011]. Bununla birlikte, uzun süreli belleğin kalıcı olup olmadığı veya yaşam sürecindeki değişikliklerin, yeni koşullara uyumun olup olmadığı sorusu açık kalmaktadır. Kesin olarak belirlenmişse, değişmez belleğin nasıl saklanabileceği hala çok net değil. hafızanın maddi temelinin, ömrü onlarca saat olan proteinler olduğu. Tüm bu sorular birlikte, ilişkisel uzun süreli belleğin moleküler oluşum ve depolama mekanizmaları, belleğin düzenlenmesi ve değiştirilmesi mekanizmaları alanındaki araştırmaları son derece alakalı hale getirir.
Son zamanlarda, fizyologlar, nispeten yakın zamanda keşfedilen yeniden birleştirme fenomenini giderek daha sık kullanıyorlar; bu, hatırlatma sırasında (belleğin yeniden etkinleştirilmesi), güçlendirilmiş uzun süreli belleğin, hatırlatma abluka koşulları altında gerçekleştirilirse kaybolabileceğini (değiştirebileceğini) açıkça gösterdi. protein sentezi [Debiec ve diğerleri, 2002; Nader ve diğerleri, 2000; Sara, 2000], yani hatırla
131
132
BALABAN
Bilgi, zorunlu olarak, yeni bir "protein belleğinin" oluşumuna bağlı olan mevcut belleğin değiştirilmesine yol açar. Yeniden konsolidasyon olgusu hala tam olarak anlaşılamamıştır ve literatürde çoğu durumda farklı deneysel kurulumlarla ilişkili olan birçok çelişkili veri vardır. Yine de, yeniden etkinleştirme sırasındaki bellek değişimi fenomeni, fizyologlara çağrışımsal süreçlerin mekanizmalarını incelemek için yeni bir yol sağlar.
Bu çalışmada, kendi ve yayınlanmış verilerimize dayanarak, doğanın mevcut hafızayı değiştirmek ve yeni koşullarda hafızanın yeniden aktivasyonu sırasında optimal adaptasyonu değiştirmek için kullandığı olası bir moleküler mekanizma hipotezini doğrulayacağız.
YENİDEN AKTİVASYON SIRASINDA BELLEĞİN LABİLİZASYONU/DEĞİŞTİRİLMESİ
Literatür, pekiştirme olmadan öğrenmenin hatırlatılmasından da kaynaklanan, yok olmaya gerçekten alternatif bir süreci ayrıntılı olarak açıklar, ancak bu durumda yeniden etkinleştirme, geçici proteine bağımlı hafıza labilizasyonu ve daha fazla konsolidasyon sürecine neden olur. İlk kez Misanin ve diğerleri [Misanin ve diğerleri, 1968] tarafından farelerin basit bir "Pavlovcu" kaçınma koşullu refleks modelinde (pavlovcu korku koşullandırma görevi) eğitildiği gösterildi . koşullu bir uyaran (hatırlatıcı) ve ardından ani bir elektrokonvülsif maruz kalma ile sunulduğunda, şoktan önce kendilerine öğretilenleri unuturlar. Hatırlatmadaki hafıza kaybı, gerçek öğrenme prosedüründen hemen sonra elektrik şoku uygulanmasındakine benzerdi. Hatırlatıcı olmadan elektrik çarpması, hafızada bu tür değişikliklere yol açmadı. Elektrik şokunun amnestik etkisinin sadece hafıza pekiştirme aşamasında değil, aynı zamanda önceden oluşturulmuş sabit bir hafızanın değişken bir duruma geçerek çeşitli amnestiklerle rahatsız edilebildiği hafızanın yeniden etkinleştirilmesi sonucunda da mümkün olduğu sonucuna varılmıştır. müdahaleler [Mactutus ve diğerleri, 1979]. 1990'ların ortalarına kadar, aslında bu fenomen hakkında hiçbir yayın yoktu, ancak daha sonra hafızanın yeniden etkinleştirilmesi sırasında neler olduğunu anlatan çalışmaların sayısı çarpıcı bir şekilde arttı. K.V.'nin laboratuvarında. Anokhin, tavuklar üzerinde, protein sentezi inhibitörlerinin intrakraniyal kullanımının, öğrenmeyi hatırlattıktan sonra yalnızca belirli bir süre sonra hafıza bozukluğuna neden olabileceği gösterilmiştir [Litvin, Apokichn, 1999]. [Nader ve diğerleri, 2000]'de, elektrik şoku hafızasının yeniden etkinleştirilmesinden hemen sonra sıçanların bazolateral amigdalasına bir protein sentezi blokerinin (PSB) infüzyonunun, tepki olarak donma tepkisinde önemli bir azalmaya neden olduğu bulunmuştur. koşullu uyarıcı Bu deneylerde öğrenmeden yeniden etkinleştirme gecikmesi 14 güne kadar olabilir, ancak BSB'nin bir hatırlatmadan sonraki etkili eylem süresi altı saati geçmedi, bu da hafızanın yeniden birleştirilmesinin zaman sınırlarını gösterir. Bu veriler, sıçanlarda farklı öğrenme paradigmaları ile farklı BSB'ler ile doğrulanmıştır [Debiec ve diğerleri, 2002; Duvarcı ve diğerleri, 2006]. Çeşitli hayvanlar üzerinde yeniden konsolidasyon fenomenini anlatan çalışmalar vardı [Eisenberg ve diğerleri, 2003; Pedreira ve Maldonado, 2003; Suzuki, 2004; Gainutdinova ve diğerleri, 2005].
Hatırlatma ve müteakip yeniden pekiştirme mevcut belleği siler mi yoksa kullanımını engeller mi sorusunun cevabı [Anckhin ve diğerleri, 2002] C. Alberini ve diğerleri tarafından çok ilginç bir çalışmada verilmektedir. [Inda ve diğerleri, 2011]. Bu çalışmada, farelerde pasif kaçınmanın (inhibitör kaçınma, deney bir elektrik şokundan sonra karanlık bir odaya girme gecikmesini ölçmektedir) geliştirilmesinden sonra geçen süreye bağlı olarak, bir hatırlatıcının hafızayı güçlendirebileceği deneysel olarak gösterilmiştir ( eğitimden iki hafta sonrasına kadar) veya ardışık testler sırasında (eğitimden dört hafta sonra) hafızayı söndürmek için. Bu yazarlara göre, yeniden konsolidasyonun protein sentezine olan bağımlılığı zamanla gizemli bir şekilde ortadan kalkmaktadır [Inda ve diğerleri, 2011].
Hafızanın yeniden birleştirilmesi, çoğu omurgalıda ve birçok omurgasızda tanımlanmış, evrimsel olarak korunmuş bir fenomendir ve bu fenomenin, hafıza oluşumu ve depolanmasının temel özelliklerini yansıttığını düşündürmektedir. Çeşitli omurgalı ve omurgasız türlerinde beyin yapısının büyük çeşitliliği, yeniden konsolidasyon olgusunu beynin sistemik bir özelliği olarak değil, daha ziyade herhangi bir hayvanda bulunabilen temel bir nöral mekanizma olarak düşünmemize izin vermez.
YÜKSEK SİNİR AKTİVİTE DERGİSİ Cilt 67 Sayı 2 2017
BELLEK MODİFİKASYONUNUN MOLEKÜLER MEKANİZMALARI
133
ancak beynin kendine özgü mimarisinden bağımsız olarak gelişmiş bir sinir sistemi.
Yeniden birleştirme paradigması, bellek modifikasyon mekanizmalarını incelemek için son derece uygundur ve sonraki bölümlerde, doğanın yalnızca belirli bir biçimden sorumlu sinir ağlarında sinaptik bağlantıların etkinliğini seçici olarak değiştirme yeteneğini nasıl sağladığını göstermeye çalışılacaktır. bellek.
BELLEK DEPOLAMA İÇİN MOLEKÜLER SİSTEM
Bellek değişiminin olası mekanizmalarını analiz etmeden önce, bellek depolamanın olası alt katmanlarını açıklamak mantıklıdır. Geriye dönüp bakıldığında, hemen hemen her nörokimyasal sistemi bloke etmeye yönelik tüm girişimlerin hafızayı ihlal ettiği ve sinir hücresinde çalışan biyokimyasal sistemlerin çoğuna sürekli olarak hafızanın temeli denildiği söylenebilir. Vakaların %100'ünde hafıza oluşturma yeteneğinin de bozulmuş olması, yani tüm bu moleküler sistemlerin hafıza depolamaya özgü olmaması son derece önemlidir.
Nispeten yakın bir zamanda, blokajı uzun süreli hafızayı bozan, ancak yeni bir hafıza oluşturma olasılığını değiştirmeyen bir hafıza molekülünün var olma olasılığına ilişkin veriler ilk kez ortaya çıktı. Sürekli aktif protein kinaz C'nin (PKC) bir izoformu olan protein kinaz Mzeta'nın (PKM Z ), rahatsız edici uzun süreli belleğin depolanması için gerekli olduğu gösterilmiştir. Özellikle, protein kinaz Mzeta'nın hipokampusta [Pastalkova ve diğerleri, 2006] uzamsal hafızayı ve neokortekste şartlı tat alma isteksizliğini depolaması ihtiyacı gösterilmiştir [Shema ve diğerleri, 2007].
Başlangıçta, Mzeta protein kinazı, uzun vadeli güçlenmeyi sürdürmek için gerekli ve yeterli olan benzersiz kinaz yapısı nedeniyle sürekli aktif olarak tanımlandı [Sacktor ve diğerleri, 1993; Ling ve diğerleri, 2002]. Çoğu PKC izoformu, ikinci haberciler için bağlanma bölgelerine sahip düzenleyici alanlar ve N-terminalinde bir oto-inhibitör psödosubstrat sekansı içerirken, C-terminalinde katalitik alanlar mevcuttur [Nishizuka, 1995]. Aktivasyonun yokluğunda, psödosubstrat katalitik bölge ile etkileşime girer ve enzim otoinhibe edilmiş bir dinlenme durumundadır. Diasilgliserol veya kalsiyum gibi ikinci haberciler, düzenleyici alanı bağlayarak tam teşekküllü PKC'yi aktive ederek otoinhibisyonu hafifleten konformasyonel değişikliklere neden olur. Protein kinaz Mzeta, düzenleyici bir alanı olmayan yalnızca bağımsız bir katalitik alana sahiptir ve bu nedenle bu enzim sürekli olarak otonom olarak aktiftir. Omurgalı beyninde, protein kinaz Mzeta, PKC geninde bulunan kendi promotörü tarafından üretilir.sadece Z katalitik alanını kodlayan PKM Z mRNA'nın transkripsiyonunu tetikleyen Z [Hernandez ve diğerleri, 2003].
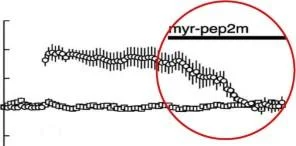
Plastik değişikliklerin indüksiyonu sırasında, tetanik stimülasyon de novo PKM Z sentezini tetikleyerek kalıcı aktif kinaz miktarını arttırır [Kelly ve diğerleri, 2007]. Bu kinazın sürekli aktivitesi, eksik PKC Z düzenleyici alanın psödosubstrat yapısını taklit eden zar geçirgen bir zeta inhibe edici peptid (ZIP) ile PKM Z aktivitesinin bloke edilmesiyle gösterildiği gibi, gelişmiş sinaptik etkinliğin sürdürülmesi için kritik öneme sahiptir . PKM Z ablukası ilehipokampustaki sinaptik güçlenme indüksiyondan bir gün sonra bile kaybolur [Pastalkova ve diğerleri, 2006]. Bu engelleyici, güçlendirme olmadan sinaptik etkinliği değiştirmediğinden, bu etki güçlendirilmiş sinapslara özgüdür.
PKM Z'nin LTP'yi ve uzun süreli belleği sürdürmedeki rolüne işaret eden kanıtların çoğu (yukarıya bakın), protein kinaz aktivitesinin ZIP tarafından farmakolojik inhibisyonuna dayanmaktadır. Bu ışıkta, uzun vadeli değişiklikleri incelemek için alternatif yaklaşımlar kullanan bazı çalışmalar, PKM Z'ye ek olarak , uzun süreli belleğin sürdürülmesinde aktif olarak yer alan aynı aileden diğer moleküllerin varlığı sorusunu gündeme getirdi .
PKM Z'nin rolünü gözden geçirmek için ana argüman, inaktive edilmiş bir PK^/PKM Z genine sahip fareler üzerinde yapılan son çalışmalarda elde edilen sonuçlardı [Lee ve diğerleri, 2013; Volk ve diğerleri, 2013; Tsokas ve diğerleri, 2016]. Paradoksal olarak, PK^/PKM Ç geninin nakavt (geleneksel nakavt) farelerde bir dizi farede ilişkisel, uzamsal ve motor belleğin depolanması ve geri alınması üzerinde hiçbir etkisi olmamıştır.
YÜKSEK SİNİR AKTİVİTE DERGİSİ Cilt 67 Sayı 2 2017
134
BALABAN
davranış testleri İn vivo veriler , bu hayvanların hipokampal bölümlerinin uyarılmasının hücrelerde PKM1 Z olmamasına rağmen LTP gelişimine yol açtığı bir dizi in vitro deneyle desteklenmiştir [Volk ve diğerleri, 2013; Tsokas ve diğerleri, 2016]. Yazarlar, ZIP varlığında kontrol ve PKC Z /PKM Z -/-bölümlerindeki uzun vadeli güçlenmenin benzer şekilde ortadan kaldırıldığını gösterdi ve bu, bu engelleyicinin başka hedefleri olduğunu gösterebilir. Bu rol için bir aday , atipik protein kinaz PKCX olan PKM Ç homologudur.
Yazarlar, ZIP , fare hipokampusunun CA1 alanındaki piramidal hücrelerde hem PKMÇ hem de PKCX tarafından indüklenen güçlenme gelişimini eşit şekilde baskıladığından, her iki atipik protein kinazın da uzun vadeli değişikliklerin sürdürülmesinde rol oynayabileceğini bulmuşlardır [Tsokas ve ark . , 2016]. Bu, PKCX'in sinaptik güçlendirmenin erken safhasında önemli olduğunu gösteren daha önceki çalışmalarla tutarlıdır [Kelly ve diğerleri, 2007; Ren ve diğerleri, 2013], LTP bakımı ise hem PKM Z hem de PKCX tarafından kontrol edilebilir [Evuarherhe ve diğerleri, 2014; Tsokas ve diğerleri, 2016].
Tsokas ve diğerleri [2016], nakavt pKc Ç /PKM Ç -/- farelerin hipokampüsünde, PKCÂ protein moleküllerinin temel miktarında bir artış gözlemlendiğini ve güçlendirilmiş hücrelerde PKCX moleküllerinin "ömrünün" de arttığını gösterdi. . Normalde, in vitro uzun süreli güçlenmenin ve in vivo uzun süreli belleğin sürdürülmesine PKM Ç aracılık eder.(yukarıya bakın), ancak yazarlar tarafından ikna edici bir şekilde gösterildiği gibi, aktif bir protein kinazın yokluğunda, hipokampal plastisitede uzun vadeli değişikliklerin sürdürülmesinde ve uzamsal belleğin korunmasında lider rol PKCX'e geçer [Tsokas ve diğerleri, 2016]. ]. Böylece, nakavt PKC Z /PKM Z -/- hayvanlarının beyninde uzun vadeli güçlenmeyi sürdürmeye yönelik temel mekanizmanın, uzun vadeli plastik yeniden düzenlemeleri sürdürmeye yönelik bağımsız olarak var olan PKCX'e bağımlı bir mekanizma ile değiştirildiği deneysel olarak doğrulandı.
Bir bellek izinin oluşumu ve korunmasının, normalde her iki atipik protein kinazın da dahil olduğu karmaşık bir süreç olduğu sonucuna varılabilir. Bazı verilere göre PKCX, LTP'nin erken evresinde postsinaptik R&L1 içeren AMPA reseptörlerinin sayısını düzenlediğinden, erken belleğin oluşumu için önemlidir. Öte yandan, PKMÇ uzun süreli güçlenme ve hafızayı sürdürmek için gerekli ve yeterlidir. Ayrıca PKMÇ'nin beyinde yetersiz ekspresyonu durumunda , hafıza izinin telafi edici, PKCX aracılı korunması devreye girer.
Bugüne kadar, yalnızca atipik protein kinazlar ailesinin protein molekülleri, protein kinazlar MZ ve CX için, varlıklarının hafızanın korunması için gerekli ve yeterli olduğu doğru bir şekilde kanıtlanmıştır [Sacktor, 2011, 2012 ; ... Tsokas ve diğerleri, 2016]. Ayrıca, PKM Z'nin inhibisyonueski anıların silinmesine neden olabilir [Kwapis ve diğerleri, 2012; Migues ve diğerleri, 2010; Shema ve diğerleri, 2009]. Bununla birlikte, hafıza mekanizmalarını incelerken, kaçınılmaz olarak kilit sorulardan biri ortaya çıkar: Bir protein molekülünün ömrü oldukça kısa bir süre ile sınırlıysa, o zaman bilgi nasıl uzun süre saklanır? Bu sorunun cevabı, ancak antrenman sonucunda değişen protein konsantrasyonunun belirli bir seviyesinde bazı aktif yerel bakım sürecinin varlığı olabilir. Eğitilmiş nöronların dendritlerinde PKM Ç protein moleküllerinin yüksek seviyesini korumak için pozitif bir geri besleme döngüsü olduğu deneysel olarak gösterilmiştir.: yeni oluşturulmuş ve yapısal olarak aktif bir protein kinaz, çeviri bloğunu kaldırarak kendi çeviri sürecini etkinleştirebilir [Sacktor, 2011].
Laboratuvarımızda üzüm salyangozu (Helix lucorum) üzerinde yapılan davranış deneylerinde , peptit bloker PKM Ç'nin eğitimden bir gün sonra uygulanmasının, salyangozun şok verildiği ortamın hafızasını bozduğu ikna edici bir şekilde gösterilmiştir . Farklı ortamlardaki hayvan tepkileri arasındaki farklar tamamen ortadan kalkar (Balaban ve diğerleri, 2015). PKM Z'nin , durumun hatırlatılmasıyla aynı anda yeniden konsolidasyonun tetiklenmesiyle bloke edilmesinin, yeniden konsolidasyon tarafından tetiklenen süreçler peptit blokerin etkisinden çok daha uzun sürdüğü ve yeni olduğu için hafızanın kaybolmasına yol açmadığına dikkat etmek önemlidir. PKM Z molekülleri sentezlenir [Balaban ve diğerleri, 2015].
İn vitro elektrofizyolojik deneylerde de kanıtlanmıştır .
YÜKSEK SİNİR AKTİVİTE DERGİSİ Cilt 67 Sayı 2 2017
BELLEK MODİFİKASYONUNUN MOLEKÜLER MEKANİZMALARI
135
PKM Z blokerinin , tetanizasyondan 90-120 dakika sonra uygulanması durumunda uzun vadeli güçlenme belirtilerini ortadan kaldırdığı [Balaban ve ark., 2015]. Elde edilen sonuçlar, omurgalılarla ilgili verilerle tamamen örtüşüyor ve hem yeniden birleştirme fenomeninin hem de bir atipik kinaz ailesi kullanılarak hafıza depolama mekanizmasının evrimsel olarak muhafazakar olduğunu ve sinir sisteminin yapısına bağlı olmadığını düşünmemize izin veriyor.
Elde edilen verileri karşılaştırarak, aşağıdaki resmi oluşturabiliriz: nöronların aktivasyonu ve hücre içi kalsiyumun büyümesi, birçok kinazın (CaMKII, PI3K, ERK, PKA ve mTOR] ve bunlarla ilişkili hücre içi kaskadların aktif durumuna yol açarak fosforilasyona neden olur. hedeflerin sayısı, bunların arasında PIN1 Fosforilasyonu, translasyon bloğunu kaldırır ve aktif nöronlardaki PKM Z proteini miktarını arttırır . De-novo sentezlenen PKM Z , PDK1 fosforilasyonuna uğrar ve yapısal olarak aktif hale gelir. Ayrıca, bu PKM Z , translasyonunu orantılı olarak arttırır. Pozitif bir geri besleme döngüsü yoluyla aktive edilmiş hücreler, böylece PKM Z protein konsantrasyonunun uzun vadeli korunmasına katkıda bulunuryeni bir seviyeye ve unutulmaz bir iz bırakarak.
BELLEK MODİFİKASYONU İÇİN MOLEKÜLER SİSTEM
Son yıllarda, nitroksidin beyin de dahil olmak üzere hemen hemen tüm organların işleyişindeki rolü hakkında büyük miktarda veri birikmiştir. Önceleri, NO sentazın omurgalılar ve omurgasızlarda sadece bazı sinir hücrelerinde büyük miktarlarda bulunduğu düşünülüyordu, ancak NO sentezini bloke etmek veya temizleyiciler kullanmak (serbest bir NO molekülünü bağlamak) hemen hemen tüm durumlarda NO'nun varlığına ihtiyaç duyulduğunu gösterdi. hafızanın oluşumu, sağlamlaştırılması ve nöronal sistemlerin plastisitesi dahil birçok süreç [Jaclet, 1997; Müller, 1997; Fedele ve Raiteri, 1999; Hawkins ve diğerleri, 1998; Gül, 2000; Schweighofer & Ferriol, 2000; Katzoff ve diğerleri, 2002; Antonov ve diğerleri, 2007]. Lipofilik NO molekülünün neredeyse tüm yaşam süreçlerine katılımının evrenselliği ve çoğu zaman birbiriyle çelişen çok büyük veri tutarsızlığı,
Hipotezi formüle etmenin ilk temeli, tüm organik kimya ders kitaplarında bulunan serbest radikal NO'nun, enzimler (guanilat siklaz) dahil olmak üzere birçok bileşikle veya diğer protein ve protein olmayanlarla kovalent bir bağa girme özelliğiydi. Bileşikler. Proteinlerle etkileşime girmenin ana yollarından biri, NO'nun sisteinin tiol grubuna kovalent bağlanması olan S-nitrosilasyondur, bu da proteinin fizyolojik işlevini değiştirir ve çoğu durumda işlevsel rolünün baskılanmasına yol açar. Guanilat siklaz sistemi yoluyla ikinci etkileşim yolu, hücre içi aracıların kaskadlarını aktive eder ve nitrozilasyona uğramış olanlar da dahil olmak üzere artan protein sentezini tetikler. Böylece aslında aynı zamanda NO proteinlerin fonksiyonlarını baskılar ve yeni proteinlerin sentezini aktive eder. NO eyleminin bu ikiliği, birçok yazara göre mevcut belleğin silindiği/bloke edildiği ve yeni bir belleğin oluşturulduğu, belleğin yeniden sağlamlaştırılması sürecindeki rolü hakkında düşünmemize yol açtı [Nader ve diğerleri, 2000; Anokhin ve diğerleri, 2002; Duvarcı ve diğerleri, 2008]. Son zamanlarda, salyangoz üzerinde nispeten basit davranışsal deneylerde hafızanın yeniden birleştirilmesi olasılığını gösterdik [Gainutdinova ve diğerleri, 2005]. Bu deneylerde, belirli bir ortamdan kaçınmak için eğitilmiş salyangozlara yeni proteinlerin sentezini engelleyen bir maddenin eklenmesi ve BSB'nin arka planına karşı bu ortamın hatırlatılmasının, test sırasında hafızanın tamamen kaybolmasına yol açtığı bulundu. günler. Bundan (1) yeni proteinlerin sentezine, yani yeni bir hafızanın oluşumuna ihtiyaç duyulduğu sonucuna varabiliriz, hatırlatıldığında, ve (2) yeniden etkinleştirme sırasında "eski" hafızanın bir istikrarsızlaştırma (ya da kararsızlaştırma ya da silme) sürecinin varlığıyla ilgili sonuca. Bu koşullar altında önceden var olan hafızanın kaybolmasına NO karışırsa, NO sentezinin bloke edilmesinin bu hafızayı koruyabileceğini varsaydık. Son deneylerde,
YÜKSEK SİNİR AKTİVİTE DERGİSİ Cilt 67 Sayı 2 2017
136
BALABAN
salyangoz polisleri [Balaban ve diğerleri, 2014], NG üretiminin bloke edilmesinin veya NG temizleyicilerinin kullanılmasının, NG'nin rolü hakkındaki hipotezi doğrulayan BSB'nin arka planına karşı yeniden etkinleştirme sırasında uzun süreli belleğin kaybolmasını önlediğini ikna edici bir şekilde gösterdi. hafıza silmede.
Literatürde, öğrenme ve hafıza analizinde NG-sentaz blokerinin diğer farmakolojik ajanlarla eş zamanlı olarak kullanıldığı sadece bir çalışma bulabildik. Bu çalışmada [Wass ve diğerleri, 2006], fensiklidinin antrenmanın yapıldığı ortamla ilgili hafızayı bozduğu ve bir NG sentaz bloker ve fensiklidinin birlikte kullanımının hafıza bozukluğuna yol açmadığı, yani bu veriler hafıza bozuklukları için NG ihtiyacı olarak yorumlanabilir. Yazarlar, çevrenin hafızasını anahtar uyaranların hafızasından ayırmadan, sadece NG üretiminin blokajının hafıza üzerindeki koruyucu etkisinden bahsediyorlar.
Literatürde, NG'nin iki yüzlü bir molekül olduğu fikri zaten var, çünkü bu molekül hem sitotoksik hem de sitoprotektif etkilere sahip ("bir Janus molekülü", bkz. [Calabrese ve diğerleri, 2009]).
Yeni hafızanın oluşumu genellikle "eski" hafızanın varlığına veya yok olmasına bağlı olarak kabul edilmez, bu nedenle bu çalışma katmanını analiz etmeyeceğiz. Bununla birlikte, bir yeniden öğrenme durumunda, "eski" hafıza ya engellenmeli ya da silinmelidir. Eski hafızanın dışavurumlarının ortadan kalkması için NG'nin katılımının gerekli olduğunu bu durumda gösterebildik. Bellek konsolidasyonu dönemi ve protein sentezi ihtiyacı hakkında bilinen verileri dikkate alarak, en basit varsayım, protein moleküllerinin işlevlerinde, yeniden konsolidasyon sırasında bir blokajın arka planına karşı davranış düzeyinde olan yerel bir değişikliği varsaymak olacaktır. yeni protein sentezi, eski hafızanın kaybolması şeklinde kendini gösterecektir. Aynı koşullar altında NG üretimi ayrıca bloke edilirse mevcut hafıza korunur [Balaban ve diğerleri, 2014]. Esasen, memeliler üzerinde aynı paradigmada deneyleri tekrarlayabildiğimizi. Bir protein sentezi blokerinin kullanımı, sıçanlarda şartlı bir uyarana (ses) göre hafızanın yeniden etkinleştirilmesi sırasında hafızada önemli bir azalmaya neden olurken, bir nitrik oksit sentez blokerinin eklenmesi bu etkiyi ortadan kaldırdı ve hafıza, kontrolden bile daha iyiydi [Bal ve et al., 2016].
Hatırlatma üzerine belleğin yeniden pekiştirilmesi olgusu doğrudan eski belleğin bloke edilmesi/silinmesi/dengesinin bozulması ve yerine yenisinin oluşturulması fikrini içermektedir [Duvarcı ve Nader, 2004; Nader ve diğerleri, 2000; Anokhin ve diğerleri, 2002; Sara, 2000], bu yüzden NG'nin bellekteki rolünü incelemek için bu yaklaşımı kullandık. Hatırlatma prosedürü sırasında NG üretiminin bloke edilmesi sırasında eski hafızanın korunmasına ilişkin elde edilen veriler, NG'nin eski hafızayı silme/bloke etme ihtiyacını doğrudan göstermektedir [Balaban ve diğerleri, 2014].
MLEKÜLER BELLEK MEKANİZMASI HİPOTEZİ MG.d^HKABHH
Bilinen verilere dayanarak, yeni bir PKM Z molekülünün oluşumunun öğrenme sürecinin bir sonucu ve bir göstergesi olduğunu varsayarak, glutamaterjik sinapsta bir hafızanın oluşumuna yol açan bir dizi olay oluşturmaya çalışalım. yeni bir hafızanın oluşumu. Her şeyden önce, aktifleştirilmiş sinapstaki plastik kaymaların indüklenmesi sırasında, glutamat reseptörleri aktive edilir (şekil) ve iyi bilinmektedir ki, nöronal nitrik oksit sentaz (nNOS) molekülü NMDA reseptörü ile de temas eden postsinaptik yoğunluk proteini (PSD95) ile doğrudan temas halinde postsinapsta bulunur, bu nedenle nNGS, birincil olarak NMDA reseptörlerinden giren kalsiyum tarafından aktive edilir [Christopherson ve Bredt, 1997; Bredt, 2003]. Ortaya çıkan küçük konsantrasyonlardaki nitrik oksit, guanilat siklaz sistemi yoluyla yeni proteinlerin sentezini aktive eder ve yerel olarak yüksek konsantrasyonlarda (güçlendirilerek), postsinapsta doğrudan kendi etrafındaki proteinleri nitrozlar. Şüphesiz, PKM Zpostsinapstaki yerleşimleri göz önüne alındığında, hafızanın yeniden etkinleştirilmesi sırasında nitrozilasyon için bir substrat da olabilir (şekil). Hatırlama sırasında nöronların öğrenme sırasındakiyle aynı yoğunlukta aktive edildiği inandırıcı bir şekilde gösterildiğinden, hafızanın geri alınmasıyla ilgili ağdaki nöronların sinapslarında nitrik oksitin yerel olarak salınması öğrenme sırasındakiyle aynı seviyede gerçekleşebilir [Gelbard-Sagiv ve diğerleri, 2008].
YÜKSEK HEPBHGH AKTİVİTE DERGİSİ cilt 67 sayı 2 2017
BELLEK MODİFİKASYONUNUN MOLEKÜLER MEKANİZMALARI
137

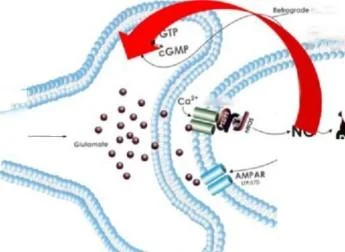
Sinapstaki plastik değişiklikler sürecinde katılımcıların ilişkilerinin şematik gösterimi. Nöronal nNOS, kalsiyumun NMDA reseptörlerinden mümkün olan en kısa sürede geçmesiyle aktive olur, çünkü nNOS, postsinaptik yoğunluk proteini PSD-95 üzerinde kollokalize edilir. Böylece, PSD-95 proteini fiziksel olarak nNOS'u NMDA reseptörlerine bağlar.
PSD-95 ile ilişkiler, nNOS'un NMDA reseptör aktivitesine bağlanmasına aracılık eder. nNOS aktivitesi birincil olarak kalmodulin ile etkileşim yoluyla nNOS'u uyaran hücre içi kalsiyumdaki Iocai artışları ile düzenlenir. Böylece PSD-95 ve ilgili proteinler, mikro yapı iskeleleri olarak işlev görür ve nNOS'u NMDA reseptörlerine fiziksel olarak bağlar.
Birleştirilmiş belleğin yeniden etkinleştirilme sürecine, proteinlerin proteazomlarda daha fazla bozunma ile poliübikitinasyona uğradığı aktif bozunmanın eşlik ettiğini not etmek oldukça ilginçtir [Hegde ve diğerleri, 2014]. Proteasomal protein degradasyonu inhibe edildiğinde hafızanın yeniden aktivasyonu sırasında BSB kullanımı ile hafıza bozukluğu olmadığından protein degradasyonu gerekli bir durumdur [Lee ve diğerleri, 2008; Jarome ve diğerleri, 2011; Ren ve diğerleri, 2013]. Protein yıkımının yalnızca yeniden birleştirme için değil, aynı zamanda uzun süreli belleği pekiştirme süreci için de gerekli olduğuna dikkat etmek önemlidir [Artinian ve diğerleri, 2008].
Son zamanlarda nitrik oksidin proteazomal protein yıkımını tetiklemede rol oynadığı gösterilmiş olup, buna duyulan ihtiyaç uzun vadeli plastik sinaptik değişikliklerde de gösterilmiştir [Bal, Balaban, 2015; Bal ve diğerleri, 2016]. Böylece, uzun süreli plastisitenin indüksiyonu sırasında bir sinir hücresinin aktivasyonu üzerine, kaçınılmaz olarak nitrik oksit oluşur ve bu, bu özel sinapsta protein yıkımı süreçlerini tetikler. Hafızanın yeniden etkinleştirilmesi ve yeniden birleştirme süreçlerinin başlatılması sırasında (inceleme için bkz. [Zyuzina, Balaban, 2015]), bu hafızanın yeniden üretilmesinde yer alan sinir ağının aktivitesinin hafıza oluşumu sırasında olduğundan daha düşük olmadığı bilinmektedir, yani. , başlatılan ve hafıza oluşumu sırasında aynı süreçler başlatılır [Gelbard-Sagiv ve diğerleri, 2008]. Anlamı,
ÇÖZÜM
Bilinen verilerin bir analizi, uzun süreli belleğin yeniden etkinleştirilmesi sırasında, bu tür belleğin altında yatan sinir ağının sinapslarında, nitrik oksidin katılımıyla, mevcut belleğin yerel bir kararsızlaştırma / istikrarsızlaştırma / silme işleminin meydana geldiğini göstermektedir. Bu durumda, ihlalleri sonraki testler sırasında hafızanın kaybolması şeklinde ortaya çıkan aynı proteinlerin sentez mekanizması başlatılır. Atipik protein kinaz M zeta, nitroksidin etkisi için bir substrat görevi görebilir, birçok hafıza türüne katılımına duyulan ihtiyaç ikna edici bir şekilde gösterilmiştir. Protein kinaz M zeta'nın aktivitesi, AMPA reseptörlerinin glutamata, postsinaptik zara verimli bir şekilde taşınması için gereklidir ve sinapsın etkinliği doğrudan AMPA reseptörlerinin miktarı ile belirlenir. Böylece,
Bu çalışma, Rus Bilim Vakfı hibe 14-2500072 tarafından desteklenmiştir.
KAYNAKÇA
Bala N.V., Balaban P.M. Ubiquitin bağımlı protein parçalanması, uzun süreli plastisite ve hafıza için gereklidir. Nörokimya. 2015. 32(4): 275-284.
YÜKSEK SİNİR AKTİVİTE DERGİSİ Cilt 67 Sayı 2 2017
138
BALABAN
Zyuzina A.B., Balaban P.M. Hafızanın yok olması ve yeniden birleştirilmesi. Yüksek Dergisi sinirli aktivite onlara. IV Pavlova. 2015 65(5): 564-576.
Anokhin KV, Tiunova AA, Rose SPR Hatırlatıcı etkileri — yeniden konsolidasyon mu yoksa geri alma açığı mı? Genç civcivlerde pasif kaçınma görevi için hatırlatmanın ardından protein sentezi inhibitörleri ile farmakolojik diseksiyon. Avro. J. Neurosci. 2002. 15(11): 1759-65.
AntonovI., HaT, Antonova I., MorozL.L., HawkinsR.D. Aplysia'da klasik sifon çekilmesinde nitrik oksidin rolü. J. Neurosci. 2007.27:10993-11002.
Artinian J., McGauran A.-MT, De JaegerX., Mouledous L., Frances B., Roullet P. Protein sentezinde olduğu gibi, protein degradasyonu, yalnızca uzun süreli uzamsal bellek konsolidasyonu değil, aynı zamanda yeniden konsolidasyon sırasında da gereklidir. Avro. J. Neurosci. 2008.27:3009-3019.
Bal N., Roshchin M., Salozhin S., Balaban P. Nitric Oxide Upregulated Proteasomal Protein Degradation in Neurons. Hücre Mol. nörobiyol. 2016. doi 10.1007/s10571-016-0413-9
Balaban PM, Roshchin M., Timoshenko AK, GainutdinovK.L., Bogodvid TK, Muranova LN, Zuzina AB, Korshunova TA Nitrik oksit, karasal salyangozlarda yeniden konsolidasyon sırasında konsolide bağlam belleğinin labilizasyonu için gereklidir. Avro. J. Neurosci. 2014. 40: 2963-2970.
Balaban PM, Roshchin M., Timoshenko AK, Zuzina AB, Lemak M., lerusalimsky VN, Aseyev NA, Malyshev AY Homolog of protein kinaz M Z , karasal salyangoz Helix'te bağlamsal caydırıcı hafızayı ve altta yatan uzun vadeli kolaylaştırmayı sürdürür. ön. hücre. nörobilim 2015.9:222.
Bredt DS Nitrik oksit sinyal özgüllüğü, sorunun kalbidir. J. Hücre. Sc.2003. 116:9-15.
Cai D., Chen S., Glanzman DL Aplysia'da uzun vadeli kolaylaştırmanın Postsinaptik düzenlemesi. Curr. Biol. 2008. 18: 920-925.
Calabrese V, Cornelius C., Rizzarelli E., Owen JB, Dinkova-Kostova AT, Butterfield DA Hücrenin hayatta kalmasında nitrik oksit: bir janus molekülü. antioksidan. redoks. sinyal. 2009. 11: 2717-2739.
Christopherson KS, Bredt DS Uyarılabilir dokularda nitrik oksit: fizyolojik roller ve hastalık. J.Clin. Yatırım. 1997. 100: 2424-2429.
Debiec J., LeDoux JE, NaderK. Hipokampusta hücresel ve sistem yeniden konsolidasyonu. Nöron. 2002. 36: 527-538.
Duvarcı S., Mamou C. Ben, Nader K. Sönme, korku anılarının bazolateral amigdalada yeniden konsolidasyona uğramasını önlemek için yeterli bir koşul değildir. Avro. J. Neurosci. 2006. 24: 249-260.
Duvarcı S., Nader K. Korku hafızasının yeniden pekiştirilmesinin karakterizasyonu. J. Neurosci. 2004. 24: 9269-9275.
Duvarcı S., Nader K., LeDoux JE De novo mRNA sentezi, amigdaladaki korku anılarının hem pekiştirilmesi hem de yeniden pekiştirilmesi için gereklidir. öğrenmek. Mem. 2008. 15: 747-755.
Eisenberg M., Kobilo T, Berman DE, Dudai Y. Geri kazanılan belleğin kararlılığı: iz baskınlığı ile ters korelasyon. Bilim. 2003. 301: 1102-1104.
Evuarherhe O., Barker GRI, Savalli G., Warburton EC, Brown MW Medial prefrontal kortekste atipik PKC inhibitörü ZIP tarafından bozulan erken bellek oluşumu, ancak hipokampusta değil. hipokampus. 2014. 24: 934-942.
Fedele E., RaiteriM. Serebral glutamat reseptörü/NO/cGMP yolunun in vivo çalışmaları. Prog. Nörobiyol. 1999. 58: 89-120.
Gainutdinova TH, Tagirova RR, Ismailova AI, Muranova LN, Samarova EI, Gainutdinov KL, Balaban PM Karasal salyangozda bağlamsal uzun süreli belleğin yeniden birleştirilmesi, protein sentezi gerektirir. öğrenmek. Mem. 2005. 12: 620-625.
Gelbard-Sagiv H., Mukamel R., Harel M., Malach R., Fried I. Serbest hatırlama sırasında insan hipokampüsündeki tek nöronların dahili olarak oluşturulmuş reaktivasyonu. Bilim. 2008. 322: 96-101.
Hawkins RD, Son H., Arancio O. Nitrik oksit, hipokampusta uzun vadeli güçlenme sırasında retrograd bir haberci olarak. Prog. beyin. Res.1998. 118:155-172.
Hegde AN, Haynes KA, Bach SV, Beckelman BC Yerel ubikuitin-proteazom aracılı proteoliz ve uzun vadeli sinaptik plastisite. ön. Mol. nörobilim 2014.7:fnmol.2014.00096.
Hernandez AI, Blace N., Crary JF, Serrano PA, Leitges M., Libien JM, Weinstein G., TcherapanovA., Sacktor TC Protein Kinaz M Sentezi From a Brain mRNA Encoding an Independent Protein Kinaz C Katalitik Alan: moleküler için çıkarımlar hafıza mekanizması. J Biol. kimya 2003. 278: 40305-40316.
Inda MC, Muravieva EV, Alberini CM Hafızanın geri alınması ve zamanın geçişi: Yeniden birleştirme ve güçlendirmeden yok olmaya. J. Neurosci. 2011. 31: 1635-1643.
Jacklet JW Nitrik oksit sinyal omurgasızları. ters çevir Neurosc.1997. 3:1-14.
Jarome TJ, Werner CT, Kwapis JL, Helmstetter FJ. Aktiviteye bağlı protein yıkımı, amigdalada korku hafızasının oluşumu ve kararlılığı için kritik öneme sahiptir. PLOS Bir. 2011. 6: e24349.
Katzoff A., Ben-Gedalya T, Susswein AJ Aplysia'da bir yiyeceğin yenmez olduğunu öğrendikten sonra çoklu hafıza süreçleri için nitrik oksit gereklidir. J. Neurosci. 2002. 22: 9581-9594.
Kelly MT, Crary JF, Sacktor TC Uzun Vadeli Güçlendirmede Çoklu Kinazlar Tarafından Protein Kinaz M Sentezinin Düzenlenmesi. J. Neurosci. 2007. 27: 3439-3444.
YÜKSEK SİNİR AKTİVİTE DERGİSİ Cilt 67 Sayı 2 2017
BELLEK MODİFİKASYONUNUN MOLEKÜLER MEKANİZMALARI
139
Kwapis JL, Jarome TJ, Gilmartin MR, HelmstetterFJ. Protein kinaz Mzeta inhibitörü ZIP'in amigdala içi infüzyonu, ön plandaki bağlam korku hafızasını bozar. nörobiyol. öğrenmek. Mem. 2012. 98: 148-153.
Lee AM, Kanter BR, WangD., Lim JP, Zou ME, Qiu C, McMahon T., Dadgar J, Fischbach-WeissS.C., MessingR.O. Prkcz boş fareler, normal öğrenme ve hafıza gösterir. Doğa. 2013. 493: 416-419.
Lee S.-H., Choi J.-H, Lee N, Lee H.-R, Kim J.-I., YuN.-K., ChoiS.-L, LeeS.-H, KimH, KaangB.- K. Sinaptik Protein Bozulması, Alınan Korku Hafızasının Dengesizleşmesinin Altında Yer Alır. Bilim. 2008. 319: 1253-1256.
LingD.SF, Benardo LS, Serrano PA, Blace N., Kelly MT, Crary JF, Sacktor TC Protein kinaz M Z , LTP bakımı için gerekli ve yeterlidir. Nat. nörobilim 2002.5:295-296.
Litvin OO, Anokhin KV Civcivlerde edinilmiş davranışsal deneyimin geri alınması sırasında hafızanın yeniden düzenlenmesi mekanizmaları: beyindeki protein sentezi blokajının etkileri. Zh. Vyssh. sinir Deiat. 1999. 49(4): 554-65.
Mactutus CF, RiccioD.C., Ferek JM Eski (yeniden etkinleştirilmiş) bellek için geriye dönük amnezi: bazı anormal özellikler. Bilim. 1979.204:13191320.
Migues PV, Hardt O., Wu DC, Gamache K., Sacktor TC, Wang YT, Nader K. PKM Z , GluR2'ye bağlı AMPA reseptör kaçakçılığını düzenleyerek anıları korur. Nat. nörobilim 2010.13:630-634.
Misanin JR, Miller RR, Lewis DJ Birleştirilmiş bir bellek izinin yeniden etkinleştirilmesinden sonra elektrokonvülsif şok tarafından üretilen geriye dönük amnezi. Bilim. 1968. 160(827): 554-555.
Müller U. Böceklerde nitrik oksit sistemi. Prog. nörobiyol. 1997. 51: 363-381.
Nader K., Schafe GE, Le Doux JE Korku anıları, geri alma sonrasında yeniden konsolidasyon için amigdalada protein sentezi gerektirir. Doğa. 2000. 406: 722-726.
Nikitin ES, Balaban PM, Kemenes G. Sinaptik olmayan plastisite, klasik koşullandırmadan sonra sinaptik etkinlikte bölümlere ayrılmış bir artışın temelini oluşturur. Curr. Biol. 2013. 23(7): 614-619.
Nishizuka Y. Sürekli hücresel tepkiler için protein kinaz C ve lipit sinyali. FASEB J. 1995. 9: 484-496.
Pastalkova E., Serrano P, Pinkhasova D., Wallace E., Fenton AA, Sacktor TC LTP'nin bakım mekanizması ile uzamsal bilginin depolanması. Bilim. 2006. 313: 1141-1144.
Pedreira ME, Maldonado H. Protein sentezi, hatırlatma süresine bağlı olarak yeniden konsolidasyona veya sönmeye hizmet eder. Nöron. 2003. 38: 863-869.
Ren S.-Q., Yan J.-Z, ZhangX.-Y, Bu Y.-F, Pan W.-W, Yao W, Tian T, Lu W PKC Â , AMPA reseptör fosforilasyonunda ve sinaptik birleşmede kritiktir. LTP. EMBO J. 2013. 32: 1365-1380.
Roberts AC, Glanzman DL Aplysia'da Öğrenme: Her iki taraftan sinaptik plastisiteye bakmak. Trendler Nörobilim. 2003. 26: 662-670.
Rose SP Tanrı'nın organizması mı? Hafıza çalışmaları için bir model sistem olarak civciv. öğrenmek. Mem. 2000. 7:1-17.
Sacktor TC PKMzeta uzun süreli belleği nasıl korur? Nat. Rev. nörobilim 2011.12:9-15.
Sacktor TC PKMzeta tarafından Bellek bakımı - evrimsel bir bakış açısı. Mol. beyin. 2012.5:31.
Sacktor TC, Osten P, Valsamis H., JiangX., NaikM.U., Sublette E. Uzun vadeli güçlenmenin sürdürülmesinde protein kinaz C'nin zeta izoformunun kalıcı aktivasyonu. Proc. Natl Acad. bilim AMERİKA BİRLEŞİK DEVLETLERİ. 1993. 90: 8342-8346.
Sara SJ Geri alma yoluyla titrek izin güçlendirilmesi. Nat. Rev. nörobilim 2000. 1:212-213.
Schweighofer N., Ferriol G. Nitrik oksidin difüzyonu serebellar öğrenmeyi kolaylaştırabilir: Bir simülasyon çalışması. Proc. Natl Acad. bilim ABD 2000. 97: 10661-10665.
Shema R., Hazvi S., Sacktor TC, Dudai Y. Neokortekste PKM tarafından hafızanın sürdürülmesi için sınır koşulları. Hafıza Öğren 2009.16:122128.
Shema R., Sacktor TC, Dudai Y. Bir PKM İnhibitörü Tarafından Korteksteki Uzun Süreli Bellek İlişkilerinin Hızlı Silme. Bilim. 2007. 317:951953.
SiK., Lindquist S., Kandel E.R. Aplysia CPEB'nin bir Nöronal izoformu prion benzeri özelliklere sahiptir. hücre. 2003. 115: 879-891.
Suzuki A. Hafızanın Yeniden Birleştirilmesi ve Sönmesi Belirgin Zamansal ve Biyokimyasal İmzalara Sahiptir. J. Neurosci. 2004. 24: 4787-4795.
Tsokas P, Hsieh C., Yao Y., Lesburgueres E., Wallace EJC, Tcherepanov A., Jothianandan D., Hartley BR, Pan L., Rivard B., Farese R..V, Sajan MP, BergoldP.J. , Hernandez A.I., CottrellJ.E, ShouvalH.Z, Fenton AA, Sacktor TC Compensation for PKM Z in long-term potentiation and uzamsal uzun-süreli bellek mutant farelerde. elife. 2016.5: doi 10.7554/eLife.14846
Volk LJ, Bachman JL, Johnson R., Yu Y., Huganir RL PKM- Z , hipokampal sinaptik plastisite, öğrenme ve hafıza için gerekli değildir. Doğa. 2013. 493: 420-423.
Wass C., Archer T., Pâlsson E., Fejgin K., Alexandersson A., Klamer D., Engel JA, Svensson L. Phencyclidine, hafızayı nitrik okside bağımlı bir şekilde etkiler: çalışma ve referans hafızası. davranış. Beyin Res. 2006. 174: 49-55.
YÜKSEK SİNİR AKTİVİTE DERGİSİ Cilt 67 Sayı 2 2017
140
BALABAN
Bellek Modifikasyonunun Moleküler Mekanizmaları
Başbakan Balaban
RAS Yüksek Sinirsel Aktivite ve Nörofizyoloji Enstitüsü, Moskova, Rusya
e-posta: pmbalaban@gmail.com
Bu incelemede, elimizdeki ve edebi verilere dayanan bir hipotez önerilmektedir. Hipotez, davranıştaki uzun vadeli değişikliklerin ve yeniden etkinleştirme nedeniyle bellekteki değişikliklerin altında yatan uzun vadeli sinaptik etkinlik değişikliklerinin moleküler mekanizmalarıyla ilgilidir. Atipik proteinkinaz M zeta'nın, ticaret yapan AMPA reseptörlerinin kontrolü yoluyla uzun vadeli sinaptik etkinlik değişikliklerindeki rolü ve bu moleküllerin, aktive edilmiş sinir hücrelerinin sinapslarında nitrik üretilen oksit tarafından nitrozilasyon olasılığı ile birlikte hipotezin temelini oluşturur. .
Anahtar Kelimeler: nöronlar, omurgasızlar, öğrenme, yeniden birleştirme, proteinkinaz mzeta.
YÜKSEK SİNİR AKTİVİTE DERGİSİ Cilt 67 Sayı 2 2017
dolambaçlı yol
- Dağın zirvesine giden yol, ister salyangoz ister dev olun, dolambaçlıdır - dedi bilge salyangoz Fujiyama'nın tepesine sürünerek
-Dağın tepesine giden yol düz değil ve salyangoz ya da dev olmanıza bağlı değil - dedi salyangoz Fujiyama'nın tepesine doğru hareket ediyor

SESSİZ, SESSİZ SÜRÜN, FUJI EĞİMİNDEN TAM YÜKSEKLİKLERE KADAR SALYANSIZ SÜRÜN
Yavaşça, yavaşça sürün, salyangoz
Fuji yokuşundan yukarı En tepeye kadar
BELLEK OLUŞUMU, DEPOLAMA VE GERİ KAZANIMININ MOLEKÜLER MEKANİZMALARI
P. Balaban
Yüksek Sinirsel Aktivite ve Nörofizyoloji Enstitüsü
Rusya Bilimler Akademisi

Hafıza?
3
Bellek, gerektiğinde kullandığımız bir "depodaki" çok fazla bilgidir -

*
Bellek, öğrenmenin sonucudur
Bellek, değişen koşullara uyum sağlamanın bir yoludur
Bellek: kısa süreli, orta, uzun süreli
Uzun süreli belleğin ana mekanizması sinaptik plastisitedir.
Sinapslar - belleğin altyapısal temeli
Hafızayı geliştiren haplar var mı?
Hafıza depoları farklı hayvanlarda aynı mıdır?
Olmayanın anısını canlı bir organizmanın beyninde silmek veya yapay olarak yaratmak mümkün müdür?
NEDEN UNUTUYORUZ?
NEYİ UNUTTUK?
6
Etkinlikteki değişiklikler hakkındaki bilgiler nasıl ortaya çıkar ve sinapslarda nasıl depolanır?
Protein sentezi olmadan yeni bir hafızanın oluşması imkansızdır.
Bir protein molekülünün ortalama ömrü - gün
Hafıza, proteinlerin katılımıyla oluşuyorsa ve yıllarca saklanıyorsa, o zaman protein konsantrasyonundaki değişiklikleri sabitlemek için bir mekanizma olmalıdır.

SİNAPTİK MEKANİZMALAR EVRİMSEL MUHAFAZACIDIR!
Sifon
musluk
+
LTP
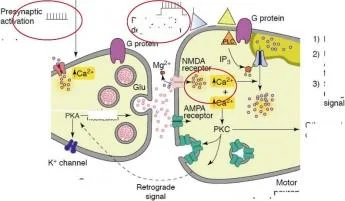
duyusal nöronlar
2) Geç faz hipereksitabilite
1) Geç faz presinaptik kolaylaştırma
vezikül mobilizasyonu
sinaptik sonrası
epolarizasyon
AMPA reseptör fonksiyonunun düzenlenmesi
Retrograd uyarımı
Diğer postsinaptik değişiklikler
nöronlar
Roberts ve Glanzman, 2003
Hafızayı geliştiren haplar var mı?
Mart 2009'da The New York Times, ciddi bir şekilde
Brooklyn'deki Tıp Merkezi'nden Dr. Sactor'un önderliğindeki bilim adamlarının, İSTİKRARIN TEMELİ'ni keşfettiklerini: insan beynindeki herhangi bir istenmeyen hafızayı kısa süre içinde silmenin mümkün olacağı bir "hafıza molekülü". sonraki hayatı boyunca onun için daha kolay. Enzim protein kinaz M-zeta, uzun süreli hafıza mekanizmasının temel unsurlarından biri olarak kabul edilir (bu birkaç yıl önce kurulmuştur), ancak yazarlara göre en önemlisi, yalnızca karmaşık anıları kurtardığı için ilginçtir. , taahhüt edilen eylemler ve deneyimler hakkında ayrıntılı bilgiler şoklar. Sonuç olarak, M-zeta protein kinaz moleküllerinin seçici yıkımı ile kişi, istenmeyen olay ve deneyimleri “unutabilir” ve beyninin işleyişi bozulmaz.
Ö'
BÖYLE
O_sh
200
250
100
150
-1 0 1 2 3 4 5 6
Zaman (saat)
hakkında
SL CO _o
250
£150
100
scr-myr-pep2m
co o_ sh
-1 0 1 2 3 4 5 6
Zaman (saat)
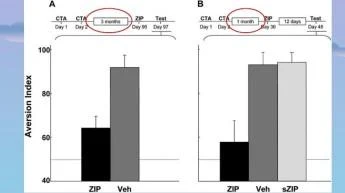
ZIP'in insular kortekste çok uzun süreli CTA belleği üzerindeki etkisi. (A) ZIP/araç, eğitimden 3 ay sonra uygulandı ve 2 gün sonra hafıza test edildi. Kesikli çizgi, CS ve su için eşit tercihi gösterir, yani AI = 50. (B) ZIP/araç/karıştırılmış ZIP, eğitimden 1 ay sonra uygulandı ve hafıza, 12 gün sonra test edildi. Sakarin, hem A hem de B'de CS idi.
PKmZ
protein kinaz
M zeta
Yapısal olarak aktif
protein kinaz C (PKC) izoformu
Hafıza depolamaya dahil
PRKCZ geni
LTP indüksiyonu
-postsinaptik
Dendritlere taşınır »
NMDAR
CaMKII, PI3K, MARK, RKA, mTOR, aktin
Çeviri bloğunu serbest bırak
AMPAR
İPİNİ PKMÇmRNA (bastırılmış bir Uy'de çeviri)
LTR'de PKMÇ oluşumu Protein kinaz C, zeta (PRKCZ) genin biri düzenleyici alanı (Reg; kırmızıyla gösterilmiştir) ve bir katalitik alanı (Cat; yeşille gösterilmiştir) kodlayan ekzonlardan tam uzunlukta bir protein kinaz CÇ (PKCÇ) üreten iki promotöre sahiptir. Nöronlarda, dahili bir promotör, düzenleyici bir alan olmaksızın bir Ç katalitik alanı kodlayan bir protein kinaz MÇ (PKMÇ) mRNA üretir. PKMÇ mRNA, dendritlere taşınır ve PIN1 (NIMA1 ile etkileşime giren protein) tarafından translasyonel olarak baskılanır. Uzun vadeli güçlendirme indüksiyonu sırasında, translasyon bloğunu serbest bırakmak için NMDAR aktivasyonu tarafından uyarılan çoklu sinyal yolları gereklidir. Önce sentezlenen PKMÇ, PKMÇ'nin konstitütif kinaz aktivitesini artıran fosfoinositide bağımlı protein kinaz 1'e (PDK1) bağlanır ve fosforile edilir.
PKMÇ daha sonra , PKMÇ mesajının artan dendritik çevirisini sürdürmek için PIN1'in inhibisyonu yoluyla bir pozitif geri besleme döngüsü başlatır. PKMÇ, kaçakçılık proteini N-etilmaleimide duyarlı faktörün (NsF) etkisiyle postsinaptik yoğunluktaki reseptörlerin sayısını artırarak AMPAR yanıtlarını güçlendirir.
CaMKII, Ca2+/kalmodulin bağımlı protein kinaz II; glu, glutamat; MAPK, mitojenle aktive olan protein kinaz; mToR, rapamisinin memeli hedefi; PI3K, fosfatidilinositol 3-kinaz; PKA, protein kinaz A
Çalışmalar, hafızanın birkaç saat - gün içinde oluştuğunu, ancak bir konsolidasyon döneminden sonra, protein moleküllerinin yerel olarak kendi kendine yeniden üretilmesiyle kalıcı olarak depolandığını göstermiştir.
Başka bir deyişle, PKMz molekülü her zaman hafızanın kritik bir bileşeni olarak kalır ve hafıza stabilitesinin temelidir.
İki farklı bağlam: 6 reklam ve gCass
BAĞLAM 1 ( TOP ÜZERİNDE )
Hafıza depoları farklı hayvanlarda aynı mıdır?
BAĞLAM 2 (CAM ÜZERİNDE)

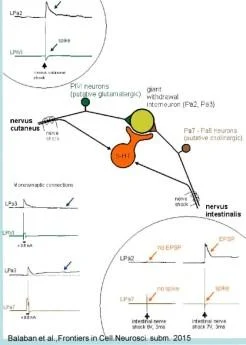
Balaban ve diğ., Frontiers in Cell.Neurosci. enjeksiyonlardan sonra 2015
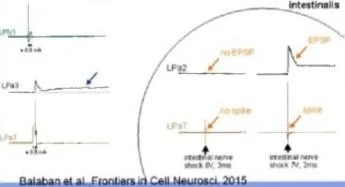
^_1Ûmin_^ PPGGP GTTP GTTGGІ
n.cutaneusj ■ !■ [■ ■ m | |
şok 1 '"
tetanoz tetanoz tetanoz
10Hz 10Hz 10Hz
UTV

ilk tetanozdan sonraki süre, min


Zaman, dakika
Balaban ve diğ., Frontiers in Cell.Neurosci. 2015
Alg Haematococcus pluvialis'in fhc fototaksisindeki fotoreseptör elektrik potansiyeli
Merhaba Yo! 271 2 Şubat 197 S
FF Litvin
OA SINESHC'HEKOV
V* A, SİNESCHHLKOV
C'hafr of Physico-chef Tlical Hiology r
Biyoloji Bölümü , Mose w Eyalet Üniversitesi
Channelrhodopsin2, tek hücreli alg Chlamydomonas reinhardtii'den izole edilen ışıkla aktive olan bir kanaldır.
Olmayanın anısını canlı bir organizmanın beyninde silmek veya yapay olarak yaratmak mümkün müdür?
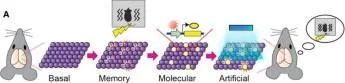
Seviye Oluşturma Etiketleme Aktivasyonu

Seviye Nötr Bellek Etiketleme
Hafif + Şok Fasle korku hafızası
Hafıza Engram Hücreleri Olgunlaştı. Neuron 87, 2 Eylül 2015 Susumu Tonegawa, Xu Liu, S Ramirez, R Redondo
Şekil 2. Hafıza Engram Hücre Popülasyonlarının Optogenetik Manipülasyonları (A) Hafıza engram hücre popülasyonunun hafif aktivasyonu, hafızanın geri çağrılmasına neden oldu. Bağlamsal bir korku hafızasının oluşumu sırasında aktif olan nöronlar, ChR2 ile etiketlendi. Bu nöronlar, farklı bir bağlamda ışık uyarımı ile yapay olarak etkinleştirildiğinde, hayvanlar, korkuyla ilişkili önceki bağlamın hatırlandığını gösteren donma davranışı sergilediler.
(B) Yanlış bir bağlamsal korku hafızasının oluşturulması. Nötr bir bağlamda aktif olan nöronlar, ChR2 ile etiketlendi ve daha sonra hayvanlara aynı anda ayak şoku verilirken farklı bir bağlamda ışıkla yeniden etkinleştirildi. Hayvanlar orijinal tarafsız bağlama geri döndüklerinde, nötr bağlam ile ayak şokunu ilişkilendiren yanlış bir hatıranın hatırlandığını gösteren korku tepkisi sergilediler. yirmi
BİLİNEN:
NO, sinaptik plastisite için gereklidir
NO, düşük konsantrasyonlarda protein sentezini aktive ederken, yüksek konsantrasyonlarda proteinleri nitrozlayarak konformasyonlarını değiştirir.
SORU:
• ? HAYIR hafıza silmeye dahil mi?

Nitrosilasyon, bir nitrozil grubunun bir proteine post-translasyonel olarak eklendiği bir protein modifikasyonudur.
Joseph Loscalzo tarafından keşfedilen S-nitrosilasyon, nitrik oksidin önemli bir biyolojik reaksiyonudur; proteinlerdeki sistein kalıntıları da dahil olmak üzere tiyol gruplarının S-nitrozotiollere (RSNO'lar) dönüştürülmesini ifade eder. S-Nitrosilasyon, ana protein sınıflarının çoğunun veya tamamının dinamik, translasyon sonrası düzenlenmesi için bir mekanizmadır.
Bellek konsolidasyonu, ilk belleğin oluşumundan hemen sonra gözlemlenen varsayımsal bir süreçtir ve belleğin oluşumunu ve stabilizasyonunu yansıtır.
Yeniden birleştirme, bellek etkinleştirildiğinde gözlemlenen belleğin yeniden dengelenme sürecidir ve birleştirilmiş bellekte değişiklik olasılığını yansıtır.

Yeniden etkinleştirme
Konsolidasyon
STM
yeniden konsolidasyon
aktif durum
OLARAK
LTM
DIR-DİR
PROTEİN SENTEZİNE BAĞLIDIR!
PROTEİN SENTEZİNE BAĞLI DEĞİLDİR!
Fid. 4 ^minder (Deney 2)
Grup (G1)
T Şoklar T1 T2
Q II II II II I ii Q RCTMer Q
♦ anizomisin
Grup2 (G2)
T Şoklar T1 T2
AllillII r ,nd 4
♦ anizomisin + L-NNA
Grup3 (G3)
T Şoklar T1 T2
II II II II II II II II II II Q Remlnjer Q
d ii - deney günleri
Hatırlamadan 20 dakika önce anizomisin/L-NNA enjeksiyonları ile bir bağlam şartlandırma deneyinin protokolü. T, T1, T2 - bağlam koşullandırma testleri.
Tig.5
^ender (Ejçperient 2)

İki farklı bağlamda ölçülen üç salyangoz grubundaki geri çekilme tepkilerinin ortalama genlikleri (±SEM): bali (güçlendirilmiş bağlam) ve cam üzerinde. Grupl (G1), n=8; Grup2 (G2), n=7; Grup3(G3), n=5.
Y ekseni - testten önceki uzunluğun %'si cinsinden dokunaç geri çekilme genliği.
***

Şok Hatırlatma + ilaçlar
Balaban ve diğ., Avrupa. J. Nörobilim, 2014
Bellek stabilitesinin temeli, yerel konsantrasyon kendi kendine bakım özelliğine sahip PKMz molekülü ve onun homologları olabilir.
Bellek plastisitesinin temeli, bellek kararlılığı proteinlerinin işlevlerini YEREL OLARAK değiştiren nitrik oksitin yerel üretimi olabilir.
NEDEN UNUTUYORUZ?
29

100
BESLENME REAKSİYONLARININ SAYISI
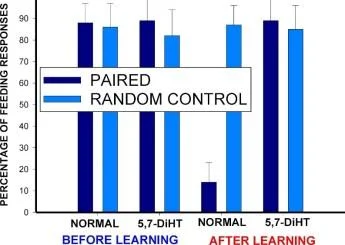
EĞİTİM SONRASI GÜÇLENDİRMEYE GEREK YOKTUR
BESLENME YANITLARININ YÜZDESİ

1 EŞLİ
1 ÇİFTLİ, 5.7-DiHT ENJEKTE

ÖĞRENMEDEN ÖNCE
ÖĞRENDİKTEN SONRA

Yeniden etkinleştirme
Konsolidasyon
yeniden konsolidasyon
STM
jrt dönemli bellek
Aktif durum"
OLARAK
LTM
DIR-DİR
PROTEİN SENTEZİNE BAĞLIDIR! GÜÇLENDİRME SİSTEMLERİNİN AKTİVİTESİ GEREKLİDİR
PROTEİN SENTEZİNE BAĞLI DEĞİLDİR!
GÜÇLENDİRME SİSTEMLERİNİN FAALİYETİNE GEREK YOK (?)
YENİDEN KONSOLİDASYON:
∙ YENİ PROTEİN SENTEZİNE İHTİYAÇ VAR
∙ BELLEĞİN GÜCÜNE BAĞLIDIR
∙ HATIRLATICI AYARLARINA BAĞLIDIR
∙ BELLEK UZUNLUĞUNA BAĞLIDIR
∙ ?GÜÇLENDİRME SİSTEMİNİN AKTİVİTESİNE BAĞLILIK?
34
35
BAĞLAM 1
BAĞLAM 2
80
- cam jej - top
QG1 -anizomisin+h atırlatıcı N=5
• G2 - 5.7 DiHT+anizomi sin+hatırlatıcı N=5
W
BÖYLE
hakkında
BÖYLE
W
W
0
W
Q tr W
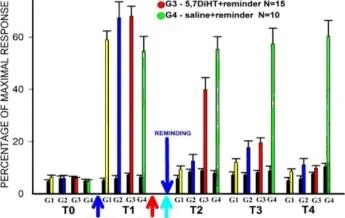
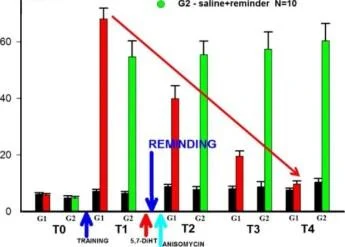
Balaban ve diğerleri, Sc. Raporlar, 2016
80
ve qlass AYAR HAFIZA D . ban O G1 -5.7DiHT+hatırlatıcı N=15

EĞİTİM
5,7-DiHT
ANİZOMİSİN
Balaban et. al., Sc. temsilci 2016
SİNYAL HAFIZASI


Balaban et. al., Sc. temsilci 2016

BESLEME SİMÜLASYONLARI
BAĞLAM UYARMALARI

MOTOR
MOTOR
PROGRAM
PROGRAM
Balaban ve diğerleri, Sc. Raporlar, 2016
sonuçlar
İlk kez, hafızayı koruma sürecini başlatmak için durumsal hafızanın yeniden aktivasyonu sırasında nöral takviye sistemini aktive etme ihtiyacı gösterilmektedir.
Anahtar bir uyaran (yiyecek) için hafıza ağının nöronları, depolanması takviye sistemine bağlı olmayan ilişkisel değişikliklere uğrar.
Takviye sistemindeki nöronal aktivitenin blokajı, (1) yeniden etkinleştirme üzerine durumsal hafızanın hızlı bir şekilde yok olmasına ve (2) durumsal hafıza bileşeninin ortadan kalkması nedeniyle bir gıda uyarıcısı için hafızanın zayıflamasına yol açar.
Beynin güçlendirici sistemlerinin katılımı olmadan hafızayı tekrar tekrar etkinleştirdiğimizde unuturuz (hafızayı değiştiririz).

dolambaçlı yol
- Dağın zirvesine giden yol, ister salyangoz ister dev olun, dolambaçlıdır - dedi bilge salyangoz Fujiyama'nın tepesine sürünerek
-Dağın tepesine giden yol düz değil ve salyangoz ya da dev olmanıza bağlı değil - dedi salyangoz Fudziyama'nın tepesine doğru hareket ediyor

SESSİZLİK. Yavaşça, yavaşça sürün, salyangoz Fudzi yokuşundan yukarı En tepeye kadar
Moleküler kararlılık mekanizmaları ve
hafıza plastisitesi
P. Balaban
Rusya Bilimler Akademisi Yüksek Sinirsel Aktivite ve Nörofizyoloji Enstitüsü


IP PAVLOV
1849-1936

Nörobiyolojinin ana sorusu, bir organizmanın bireysel gelişimi sırasında 10 11 nöronun , işlevsel bir düşünen beyin oluşturan 10 15 spesifik sinaptik bağlantıyı nasıl kurduğudur.
"kritik dönemler"
Biz hatırladığımız şeyiz... Ch.Aitmatov

Nasıl ortaya çıkıyor ve sinapslardaki aktivite değişiklikleri hakkındaki bilgiler nerede saklanıyor?
PLASTİKİYE KARŞI KARARLILIK
Protein sentezi olmadan yeni bir hafızanın oluşması imkansızdır.
Bir protein molekülünün ortalama ömrü - gün
Hafıza, proteinlerin katılımıyla oluşuyorsa ve yıllarca saklanıyorsa, o zaman protein konsantrasyonundaki değişiklikleri sabitlemek için bir mekanizma olmalıdır.

Sinaptik plastisite mekanizmaları
Sifon
musluk
CS
TAI şoku
BİZ

+
ltd.
o°S
AMPA reseptörü I
PKC
almak
2)
3)
K + kanalı
duyusal nöronlar
Retrograd sinyal
Pre X7 ..„ r etkinleştirme
1) Geç faz presinaptik kolaylaştırma
2) Geç faz hipereksitabilite
oh oh ° oh ° oh
Araç seferberliği _
sinaptik sonrası
patlama
o o o °
LTP
Glu
Mg2 +
°« hakkında?
oo
o o°
■ h o o
Ah Ah
NMDA
IP
« o°<? ° О °о2) о сЄоо oo
AMPA reseptör fonksiyonunun düzenlenmesi
Retrograd sinyalin uyarılması
Diğer postsinaptik değişiklikler
motor nöronlar
Roberts ve Glanzman, 2003
BELLEK MOLEKÜLÜ?
Mart 2009'da The New York Times, Brooklyn Tıp Merkezi'ndeki bilim adamlarının Dr.'nin hayatını kolaylaştırdığını ciddi bir şekilde duyurdu.
Enzim protein kinaz M-zeta, uzun süreli hafıza mekanizmasının temel unsurlarından biri olarak kabul edilir (bu birkaç yıl önce kurulmuştur), ancak yazarlara göre en önemlisi, yalnızca karmaşık anıları kurtardığı için ilginçtir. , taahhüt edilen eylemler ve deneyimler hakkında ayrıntılı bilgiler şoklar. Sonuç olarak, M-zeta protein kinaz moleküllerinin seçici yıkımı ile kişi, istenmeyen olay ve deneyimleri “unutabilir” ve beyninin işleyişi bozulmaz.

ZIP'in insular kortekste çok uzun süreli CTA belleği üzerindeki etkisi. (A) ZIP/araç, eğitimden 3 ay sonra uygulandı ve 2 gün sonra hafıza test edildi. Kesikli çizgi, CS ve su için eşit tercihi gösterir, yani AI = 50. (B) ZIP/araç/karıştırılmış ZIP, eğitimden 1 ay sonra uygulandı ve hafıza, 12 gün sonra test edildi. Sakarin, hem A hem de B'de CS idi.
PKmÇ
Protein kinaz M zeta
Protein kinaz C'nin (PKC) yapısal olarak aktif izoformu
Hafıza depolamaya dahil

LTP'de PKMZ oluşumu. Protein kinaz C, zeta (PRKCZ) genin iki promotörü vardır; biri, bir düzenleyici alanı (Reg; kırmızıyla gösterilmiştir) ve bir katalitik alanı (Cat; yeşille gösterilmiştir) kodlayan ekzonlardan tam uzunlukta bir protein kinaz CZ (PKCZ) üretir. Nöronlarda, dahili bir promotör, düzenleyici bir alan olmaksızın bir Z katalitik alanını kodlayan bir protein kinaz MZ (PKMZ) mRNA üretir. PKMZ mRNA, dendritlere taşınır ve PIN1 (NIMA1 ile etkileşime giren protein) tarafından translasyonel olarak bastırılır. Uzun vadeli güçlendirme indüksiyonu sırasında, translasyon bloğunu serbest bırakmak için NMDAR aktivasyonu tarafından uyarılan çoklu sinyal yolları gereklidir. PKMZ sentezlendikten sonra, PKMZ'nin yapısal kinaz aktivitesini artıran fosfoinositide bağımlı protein kinaz 1'e (PDK1) bağlanır ve fosforile edilir. PKMZ daha sonra pozitif bir geri besleme döngüsü başlatır.PKMZ mesajının artan dendritik çevirisini sürdürmek için PIN1'in inhibisyonu yoluyla . PKMZ, kaçakçılık proteini N-etilmaleimide duyarlı faktörün (NsF) etkisiyle postsinaptik yoğunluktaki reseptörlerin sayısını artırarak AMPAR yanıtlarını güçlendirir.
CaMKII, Ca2+/kalmodulin bağımlı protein kinaz II; glu, glutamat; MAPK, mitojenle aktive olan protein kinaz; mToR, rapamisinin memeli hedefi; PI3K, fosfatidilinositol 3-kinaz; PKA, protein kinaz A
M =PSD-95_=en yakın arkadaşım
* = RCMC
X = GluR2

I PSD-95 agregasyonu I postsinaptik GluR2 1 Mantar Dikenleri ?
t PSD-95 agregasyonu t postsinaptik GluR2 t Mantar Dikenleri ?
Şekil 1. PKMÇ Aktivitesinin Artış ve Azalışlarına Omurga Morfolojisi Modeli
Çalışmalar, hafızanın birkaç saat - gün içinde oluştuğunu, ancak bir konsolidasyon döneminden sonra, protein moleküllerinin yerel olarak kendi kendine yeniden üretilmesiyle kalıcı olarak depolandığını göstermiştir.
Başka bir deyişle, PKMz molekülü her zaman hafızanın kritik bir bileşeni olarak kalır ve hafıza stabilitesinin temelidir.
PLASTİKİYE KARŞI KARARLILIK
∙ Bellek konsolidasyonu, ilk belleğin oluşumundan hemen sonra gözlemlenen varsayımsal bir süreçtir ve belleğin oluşumunu ve stabilizasyonunu yansıtır.
∙ Yeniden konsolidasyon - birleştirilmiş hafızayı değiştirme olasılığının hatırlatılması ve yansıtılması sırasında gözlemlenen hafıza istikrarsızlaştırma süreci
TidL
<İki farklı bağlam: 6 adandgCass
BAĞLAM 1 ( TOP ÜZERİNDE )
BAĞLAM 2 (CAM ÜZERİNDE)
A
PROTOKOL1
günler
Bir izomisin/salin

ani
Sal
Bağlam koşullandırması için test edildikten sonra anizomisin/tuzlu su enjeksiyonu ile bağlam koşullandırma deneyi (A) protokolü, hatırlatma yok. B - iki farklı bağlamda ölçülen üç salyangoz grubundaki geri çekilme yanıtlarının ortalama genlikleri (+SEM).
Öğrenme ve Hafıza 2005
PROTOKOL 2
Hatırlatma

Bir nisomisin/salin
Hatırlattıktan hemen sonra anizomisin/tuzlu su enjeksiyonu ile bağlam şartlandırma deneyinin (A) protokolü. B - iki farklı bağlamda ölçülen üç salyangoz grubundaki geri çekilme tepkilerinin ortalama genlikleri (±SEM).
BELLEK
SİLİNDİ???
ani
al
PkMZ'nin immünokimyasal p lokalizasyonu

LRV
Meyve sineği
ІНІіх
I Lymnaea İAp2ysia
ІНІіх İlymnaea | Aplysia İLottia | Drosophi la
|LynInna ea |Aplysia İLottia
Drosophi la Battus Homo konsensü
ben Lymnaea
Aplysia
Meyve sineği
Rattus
Homo
fikir birliği
Lymnaea
Aplysia Lottia
Meyve sineği
Rattus
il
alipides, V i i i ben ben ■■ g.
SVDWWALGVLMFEMLAGRSPFD7VGNADNPDQNTEDYLFQ ben
RSPFDLAGASZNPDQNTEDYLFQiV RSPFD II - -TDNP DM NTEDYL FQV RSPFD II--TDNP Dt-' NTE DY L FQ V
500 510
A
A
N
N
D
D
□70
AERLGCH PQTGFSD IQ S
KİMLİK W IM
620
b VV » L
K a r b o k s i t e r m i n a l a l a n ı
6 3 0 6 4 0
AATNY
DED
N DQ
|N DQ
kinaz alanı
330 340
İKKALV TDDED
VKKELVHDDED
IDW VQ
IDW VQ
IDW VQ
IDW VQ
IDW VQ
IDWVQ.
IDWVQ TEKHVFE,
KG FLNK KGFLNK KG FLN için KGFLNK KGFLNK KG F LN için KGFLNK
G
İle
400
DLM ve HMQRQR DLMFHMQRQR DLM FHMQRQR DLMFHMQRQR DLM ve HMQRQR
DLM F HMQ RQ R DLMFHMQRQR
PFLVG1HSCFQT
PFL VG
PFLVG
PFLVG
PFLVG
PFLVG
PFLVG
PFLVG
LHSC FQ TLHSC FQ T LHSCFQ T LHSCFMT LHSC FQ TLHSC FQ TLHSC FQ T
Şoklar
Hatırlatma
'1 ben K2

BAĞLAM 1 ( TOP ÜZERİNDE )
T, T1, T2 - czeczi testi - deney günleri
İLAÇLAR

BAĞLAM 2 (CAM ÜZERİNDE)

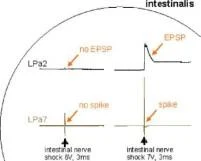
Balaban ve diğ., Frontiers in Cell.Neurosci. subm. 2015
Normalleştirilmiş EPSP genliği O
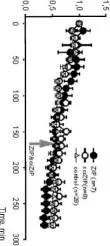
Normalleştirilmiş EPSP genliği

Ö
3
Ö
S
EPSP

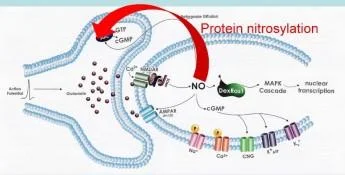
Nitrosilasyon, bir nitrozil grubunun olduğu bir protein modifikasyonudur .
post-translasyonel olarak bir proteine eklenir.

Joseph Loscalzo tarafından keşfedilen S-nitrosilasyon, nitrik oksidin önemli bir biyolojik reaksiyonudur; proteinlerdeki sistein kalıntıları da dahil olmak üzere tiyol gruplarının S- nitrozotiollere (RSNO'lar) dönüştürülmesini ifade eder . S-Nitrosilasyon dinamik bir mekanizmadır,
ana protein sınıflarının çoğunun veya tamamının çeviri sonrası düzenlenmesi.
∙ BİLİNEN:
∙ NO, sinaptik plastisite için gereklidir
∙ NO, düşük konsantrasyonlarda protein sentezini aktive ederken, yüksek konsantrasyonlarda proteinleri nitrozlayarak konformasyonlarını değiştirir.
∙ SORU:
• ? HAYIR hafıza silmeye dahil mi?
Fid. 4 ^minder (Deney 2)
Grup (G1)
T Şoklar T1 T2
Q II II II II II II Q ~
♦ anizomisin
Grup2 (G2)
T Şoklar T1 T2
Q r ,nd 4
♦ anizomisin + L-NNA
Grup3 (G3)
T Şoklar T1 2
FINN I
[=^■=1 - deneme günü
Hatırlamadan 20 dakika önce anizomisin/L-NNA enjeksiyonları ile bir bağlam şartlandırma deneyinin protokolü. T, T1, T2 - bağlam koşullandırma testleri.
(Rgminder (2 numaralı temel)
İki farklı bağlamda ölçülen üç salyangoz grubundaki geri çekilme tepkilerinin ortalama genlikleri (±SEM): bali (güçlendirilmiş bağlam) ve cam üzerinde. Grupl (G1), n=8; Grup2 (G2), n=7; Grup3(G3), n=5.
Y ekseni - testten önceki uzunluğun %'si cinsinden dokunaç geri çekilme genliği.
***
%
80-
bali cam
Balaban ve diğ., Avrupa. J. Nörobilim, 2014

Balaban ve diğ., Avrupa. J. Nörobilim, 2014
• Nitroksit sentezinin engellenmesi, yeniden konsolidasyonu engeller -"eski" bellek silinmez Olası mekanizmalar:
• "Yeni" belleğin blokajı? - Değil! (bloke protein sentezi)
• "eski" hafıza korunuyor mu? - Evet! (Protein nitrozilasyonu engellendi)
Fid.2 (Rgkarning (Ejçperiment 1)
Grup (G1)
T Şoklar T1 Besleme T2
SHIN ben
* II ■ ben
■ ■ II .
zil sesi
Grup2 (G2)
T Şoklar T1 Besleme T2
NSHI İH Q
kapamak
Zil L-NNA
Grup3 (G3)
T Şoklar T1 Besleme T2
♦ ilil hu Q
♦♦♦♦♦ ♦♦♦
L-NNA
L-NNA
t=:er=ı - deneme günleri
L-NNA/salin enjeksiyonları ile bağlam şartlandırma deneyi protokolü. T, T1,T2 - bağlam koşullandırma testleri.
Tid.Z
^ eSeagpipd (Ejçperiment 1)
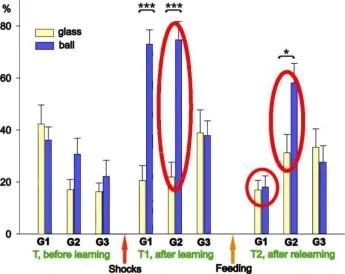
İki farklı bağlamda ölçülen üç salyangoz grubundaki geri çekilme tepkilerinin ortalama genlikleri (±SEM): bali (güçlendirilmiş bağlam) ve cam üzerinde. Grupl (G1), n=9; Grup2 (G2), n=5, Grup3 (G3), n=8.
Y ekseni - testten önceki uzunluğun %'si cinsinden dokunaç geri çekilme genliği.
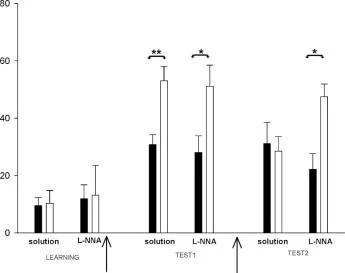
öğrenme yeniden öğrenme

EĞİTİM
Sıçanlarda
Test 1 - ANTRENMAN SONRASI
Test 2 - YENİDEN EĞİTİMDEN SONRA
■ keşif sesi
grup kontrolü (çözüm) n=8
Grup2 deneyi (l-NNA) n=6
Bellek stabilitesinin temeli, yerel konsantrasyon kendi kendine bakım özelliğine sahip PKMz molekülü ve onun homologları olabilir.
Bellek plastisitesinin temeli, bellek kararlılığı proteinlerinin işlevlerini YEREL OLARAK değiştiren nitrik oksitin yerel üretimi olabilir.
İLGİNİZ İÇİN TEŞEKKÜR EDERİZ!

Not: Bazen Büyük Dosyaları tarayıcı açmayabilir...İndirerek okumaya Çalışınız.
Yorumlar
Yorum Gönder