
Bellek, Damgalama Ve Beyin
Mekanizma araştırması
Dr. Biol tarafından İngilizce'den çeviri. Bilim O. S. VINOGRADOVA
Bellek, damgalama ve beyin. Mekanizmaların incelenmesi: Per. İngilizceden. - M .: Mir, 1988. - 343 s.,
Ünlü İngiliz nörofizyologun monografisi, damgalama mekanizmalarıyla ilgili verilerin kapsamlı bir genellemesidir . Bu davranış biçimi örnek alınarak , hafıza ve öğrenmenin yapısal ve fizyolojik temelleri analiz edilir. Kitap , yazarın Rusça baskısı için hazırladığı yeni materyalleri içeriyor.
Fizyologlar ve biyokimyacılar için - daha yüksek sinirsel aktivite uzmanları , psikologlar, etologlar.
Bu kitap ilk olarak Oxford University Press, Oxford, İngiltere tarafından İngilizce dilinde yayınlanmıştır.
ÇEVİRMEN ÖNSÖZ
Bu kitabın yazarı G. Horn'un adı ülkemizde öğrenme ve hafıza süreçleri üzerine çalışanlar tarafından çok iyi bilinmektedir. Sinir sisteminin plastisitesi ile ilgili sempozyumlara katılarak defalarca ülkemize konuk olmuştur. Bu konuyla ilgili Sovyet yayınlarında birkaç makalesi yayınlandı. G. Horn'un yazdığı bu kitap , yazar ve işbirlikçilerinin , yaşamın ilk günlerinde bazı hayvanlarda gözlemlenen özel bir davranış biçimi modeli - damgalama (imprinting) üzerindeki bellek mekanizmalarının incelenmesi konusundaki yirmi yıllık çalışmalarının sonuçlarını özetlemektedir. .
Baskı fenomenolojisi, 1930'larda K. Lorenz tarafından incelendi, ancak baskının var olduğu gerçeği geçen yüzyılda zaten biliniyordu. Bununla birlikte, bu kitabın yazarının uzun yıllara dayanan sistematik araştırmaları sayesinde, bir bellek modeli olarak damgalama titiz bir bilimsel analizin konusu haline geldi.
G. Horn'un kitabı farklı açılardan ilgi çekicidir . Her şeyden önce, biyokimya, histoloji ve elektron mikroskobundan tek sinir hücrelerinin nörofizyolojisine ve yıkım sırasındaki davranış çalışmalarına kadar beyin bilimlerinin tüm modern cephaneliğinin bir sorunu çözmek için kullanılması çok disiplinli bir yaklaşımın bir örneğidir. beyin bölgelerinin uyarılmasının yanı sıra çeşitli biyolojik olarak aktif maddeler beyne verildiğinde veya beyinden çıkarıldığında. Tarif edilen metodolojik yaklaşımların ve deneylerin hiçbiri kendi başına "kritik" değildir, ancak mantıksal bir dizi çalışma, sinir sistemine bilgi damgalamanın genellikle alışılmadık ve beklenmedik ilkelerini ikna edici bir şekilde ortaya koymaktadır.
Bu kitap başka bir açıdan öğretici. Baskılama modeli , G. Horn tarafından hafızanın en basit, en net ve en kolay ölçülebilir tezahürlerinden biri olarak seçildi. Bununla birlikte, kitap boyunca yazar, bu durumda bile beynin çalışmasında incelenmekte olan hafıza süreçleriyle tam olarak neyin ilişkili olduğunu izole etmenin ne kadar zor olduğunu ısrarla göstermektedir. 60'larda ve 70'lerde nasıl olduğunu çok iyi hatırlıyoruz. Davranışın karmaşıklığı ve beynin karmaşıklığı dikkate alınmadan biyokimyasal ve davranışsal veriler arasındaki yüzeysel korelasyonlar, belleğin doğası hakkında yanlış sonuçlara yol açtı. G. Horn'un kitabı bu tür eksikliklerden tamamen arınmış. İlk çalışma hipotezini doğrulayan sonuçlar elde edildiğinde bile (örneğin, etiketli lizin ve urasilin damgalamanın geliştirildiği tavukların beyninin sınırlı bölgelerine dahil edilmesinde bir artış), yazar bunun daha yüksek bir sonucun olabileceğini gösteriyor. motor aktivite, uyanıklık düzeyi , stres, hormon salgılanması, nöronal büyüme vb. Bu alternatif açıklamaları kademeli olarak ortadan kaldırmak ve bilgi sabitlemenin önde gelen önemini ortaya çıkarmak için yazar tarafından tanıtılan çok sayıda hayvan kontrol grubu ve deneysel etkilerin varyantları, buna bir örnektir. çeşitli bilimlerin kavşağında yatan problemlerin incelenmesinin nasıl yapılandırılması gerektiği. Hafıza izlerinin oluşumunu ölçmeyi ve çeşitli biçimlerini ayırt etmeyi amaçlayan bir dizi ustaca ve orijinal yöntemin esnek kullanımına özel önem verilmelidir - örneğin, damgalama ve "çağrışımsal öğrenme ", yani edimsel koşullu reflekslerin gelişimi. .
Yazarın uyguladığı incelikli analitik yöntemler, damgalama gibi nispeten basit bir hafıza olgusunun bile beyinde hiçbir şekilde basit bir organizasyonu olmadığını göstermeyi mümkün kılıyor . Okuyucu, damgalama ile kişinin kendi türünden bireyleri tanımaya yönelik doğuştan gelen yatkınlığı arasındaki farkı görecektir ; nesneleri damgalarken tavuk beyninin sağ ve sol yarısının işleyişindeki farklılıklar; norepinefrin ve testosteronun damgalanması üzerindeki farklı etki ve çok daha fazlası. Yazar, bu analitik verilerin sentezine dayanarak, bir bellek biçimi olarak damgalama mekanizmasının mantıksal olarak bütünsel bir konseptini sunar.
G. Horn damgalamaya ek olarak, diğer öğrenme biçimlerine de değiniyor: alışkanlık, maymunlardaki karmaşık öğrenme biçimlerinin nöronal bağıntıları. Yazarın, Batı ülkelerinde omurgalıların ve omurgasızların sinir sistemindeki plastik değişikliklerin araştırılmasında öncü olduğu belirtilmelidir ; Bu çalışmaların sonuçları, sonraki yıllarda diğer yazarlar tarafından plastik süreçleri açıklamak için önerilen birçok hipotezi öngörmüştür.
Kitapta sunulan deneysel materyal , sinapsların etkinliğindeki değişikliklerin iyonik mekanizmalarından primatlarda ve beyin hasarı olan insanlarda çeşitli bellek türlerinin saptanmasına kadar modern beyin plastisitesi kavramlarının geniş bir panoramasının üzerine bindirilmiştir .
G. Horn'un monografisi, davranışı inceleyenlere davranışın nöronal temellerini dikkate almanın ne kadar önemli ve ilginç olduğunu ve beyni inceleyenlere dikkatlice planlanmış davranışsal deneylerin önemini gösteriyor. Beyin biliminin çeşitli dallarının nihai amacının davranış mekanizmalarını aydınlatmak olduğunu hatırlıyor. Aynı zamanda basit ve anlaşılır bir dille yazılmış bu kitap, beyin aktivitesinin belirli yönleriyle yeni ilgilenmeye başlayan araştırmacılara, sırlarını ortaya çıkarmak için ne kadar heyecan verici ve sıkı bir çalışma gerektiğini gösterebilir.
OS Vinogradova
YAZARIN RUSÇA BASKIYA ÖNSÖZÜ
Davranışsal düzenlemenin sinirsel mekanizmalarının incelenmesi, SSCB'de uzun ve şanlı bir tarihe sahiptir. I. M. Sechenov, I. P. Pavlov, I. S. Beritashvili, A. R. Luria'nın çalışmaları, bu sorunun yalnızca Sovyetler Birliği'nde değil, aynı zamanda bu tür araştırmaların yapıldığı dünyanın diğer tüm ülkelerinde de derin bir etkiye sahipti. Bu bilim adamları, şu anda nöral işlevi incelemek ve davranışı analiz etmek için kullanılan oldukça verimli ve çeşitli yöntemlere sahip değillerdi . Ancak bu kadar net bir şekilde formüle ettikleri sorunları çözmek için hangi yeni ve mükemmel yöntemlerin kullanıldığını bilselerdi, şüphesiz gurur duyarlardı.
ve hafızanın nöral mekanizmalarının aydınlatılmasıdır . Bu problem ya da daha doğrusu, merkezi sinir sisteminin öğrenme süreci yoluyla modifikasyonunun açıklanmasıyla ilgili bu problemler dizisi, nörobilimin merkezinde yer alır. Aynı zamanda, bu problemler dizisi davranışı anlamak için de merkezi bir öneme sahiptir, çünkü bir bireyin - bir hayvanın veya bir insanın - varoluşunun zaman içinde sürekliliğini yaratan hafızadır. Hafıza, birbirini izleyen duyusal deneyim bölümlerini organize eden ve bir arada tutan ipliktir ve eğer bu iplik koparsa, Alzheimer hastalığında olduğu gibi hafızanın işlevi bozulursa, o zaman hafızanın bireyin korunmasındaki önemi trajik bir şekilde ortaya çıkar.
XII.Yüzyıl Fransız filozofunun ünlü sözünü yorumlamak için. Chartres'li Bernard, şunu söyleyebilirim ki, öğrenmenin sinirsel mekanizmalarına biraz daha derinlemesine nüfuz edebilseydim veüniversiteler. Çalışmam ve iş arkadaşlarımın çalışmaları hakkındaki bu rapora artık Sovyet meslektaşlarımızın kolayca erişebilecekleri için mutluyum . J
O zamandan beri, yeni damgalama deneyleri gerçekleştirildi ve bu kitapta ele alınan sorunlarla doğrudan ilgilenen çok sayıda yeni çalışma basılı olarak ortaya çıktı. Rusça yayınlarken, bu verileri metne dahil etmek için bana verilen fırsattan yararlandım.
Bu kitabın yapıcı eleştirisi ve çevirisi için arkadaşım ve meslektaşım OS Vinogradova'ya derinden minnettarım.
Cambridge, 1 Mart 1987 G. Horn
ÖNSÖZ
Öğrenme, bilginin girişi, depolanması ve çoğaltılmasını içeren karmaşık bir süreçler dizisidir. İlk öğrenme süreci farklı seviyelerde incelenir ve duyusal bilgilerin kodlanması, iletilmesi ve işlenmesi, davranışsal aktivasyon ve dikkati kapsar. Bu kurucu süreçlerin listesi tam olmaktan uzaktır. Bazılarının analizi son zamanlarda çok çarpıcı sonuçlar verdi. Aynı zamanda, üreme sürecinin fizyolojik temelleri hakkında çok az şey biliniyor ve beyindeki bilgileri sabitleme mekanizmalarına ilişkin anlayışımız yeni şekillenmeye başlıyor ve hala çok sınırlı. Bu kitabın amacı, bu alandaki bazı ilerlemeleri bildirmektir. Konunun tam bir özeti olarak değerlendirilmemeli ; bu, son yirmi yıldır dahil olduğum alışkanlık ve damgalama araştırmaları üzerine çalışmanın yalnızca bir açıklamasıdır . Bu dönemde, bir tür öğrenme olan yaygın bir davranış değişikliği biçimi olan alışkanlığı analiz etmede bazı ilerlemeler kaydettik. Bu çalışmanın bir kısmı, özellikle sinir sisteminde bilgi depolamanın doğası ile ilgili, birinci bölümde anlatılıyor. Bununla birlikte, kitabın ana bölümü baskıya ayrılmıştır. Psikoloji ders kitaplarının bu konuya çok az yer verdiğini varsayarsak, damgalama da çok özel bir davranış biçimi olarak düşünülebilir . Umuyorum ki bu kitap size damgalamanın diğer öğrenme biçimlerinden ne kadar farklı ve benzer olduğunu gösterecektir.
Hafızanın nöronal temelleri sorunu, kısmen beyin bilimleri ile davranış bilimleri arasındaki sınır çizgisinde yer aldığı için, uzun süredir araştırmalara meydan okuyor. Başarılı bir çözüm için, her iki bilim alanının temsilcilerinin entelektüel ve metodolojik yeteneklerini birleştirmenin gerekli olduğuna inanıyorum. Bu şekilde damgalama çalışmasında bir miktar başarı elde ettik. Çalışma 1966'da başladı. Bu süre zarfında, alışkanlık analizimde Pat Bateson ile tanıştım. O davranış çalışmalarında uzmandı ve ben nörofizyoloji alanındaydım. İkimizin de genel olarak öğrenmeye ve özel olarak damgalamaya ilgi duyduğumuzu fark ederek, birlikte deneysel çalışmaya başladık. Bu işbirliği çok verimli oldu.
Bu kitapta açıklanan damgalama deneylerinde birçok farklı yöntem kullanılmıştır. Sonuç olarak, kapsanan konu yelpazesi moleküler biyoloji, nörofizyoloji, nöroanatomi ve endokrinolojiden davranış incelemelerine kadar çok genişti. İnsanlar ve diğer primatlar üzerindeki çalışmaların dikkate alınması gerektiğinden, bu aralık daha da genişletilir. Listelenen alanlardan birinde çok az eğitimi olan herkesin tartışmayı kolayca takip edebilmesi için metin boyunca kullanılan terimleri açıklığa kavuşturmaya çalıştım . Kuş beyninin yapısı nispeten az kişi tarafından bilindiği için, kitabın ekinde açıklaması verilmiştir.
Hafızanın nöral mekanizmalarının ve tabii ki damgalamanın analizi tek bir "kritik" deneyde gerçekleştirilemez . Sinir sisteminde öğrenmeyle ilgili herhangi bir değişikliği belirledikten sonra, bu değişikliğin davranışın hangi yönüyle ilgili olduğunu belirlemek gerekir. Bu soru çok önemlidir. Bir deneycinin, örneğin beyindeki bazı biyokimyasal değişikliklerin öğrenmeyle ilişkili olduğundan ne ölçüde emin olduğu, bu ilişkinin özgüllüğünü kanıtlama yeteneğine bağlıdır. Ancak bu güvenilir bir şekilde kanıtlanırsa, bir sonraki, daha derin analiz düzeyine geçilebilir. Bu tam olarak çalışanlarımla birlikte izlediğim yol, bu kitabın konusu da bu. Deneysel analizin sonuçlarını yorumlamaya veya bir araya getirmeye çalışarak mantıksal akıl yürütme ihtiyacıyla yetinmedim. Bilimin daha fazla gelişmesinin kritik olarak böyle bir mantıksal senteze bağlı olduğunu düşünüyorum. Bununla birlikte, güvenilir deneysel veriler ile onlar hakkında akıl yürütme arasında ayrım yapmak çok önemlidir . Bazı bölümlerde mantıksal analizi ayrı bir kısma havale ederek aralarındaki sınırı vurgulamaya çalıştım . Bu, tüm bölümlerde yapılamadı, ancak umarım okuyucu, deneysel verilerin sağlam zeminini nerede bıraktığımı anlayacaktır.
katılan en yakın çalışma arkadaşlarıma ve eleştirileriyle kitabın metninin gelişmesine katkıda bulunan meslektaşlarıma ek olarak, tüm zorluklara rağmen taslağı yeniden daktilo eden Chris Percival'e ve Robert Hynd'a şükranlarımı sunmalıyım. Metnin tamamını okuyan, detaylı analizlere tabi tutan ve hem bana hem de kitabı eleştirel bir şekilde okuyan, metin ve kapak çizimlerini yapan ve birkaç yıl boyunca katlanan eşim Priscilla Barrett'a sürekli destek olan. nadiren hafıza , damgalama ve beyin tartışmasının ötesine geçen bir konuşma .
Ocak 1985
Cambridge GX _
BÖLÜM 1
BELLEĞİN NÖRONAL TEMELLERİNİN ANALİZİNE YAKLAŞIMLAR
Beyinde bilgi kaydetme mekanizmalarına olan ilgi uzun bir geçmişe sahiptir. Geçtiğimiz yüzyıllar boyunca ortaya atılan açıklamalar çeşitlilik gösterse de, birkaç istisna dışında bu açıklamalar ortak bir düşünceye dayanıyordu. Belirli bir deneyim ya da olay, beyindeki belirli yolların etkinliğinin oluşmasına ya da artmasına yol açar. Bu şekilde oluşan veya güçlenen bir yol , deneyimi veya olayı "temsil eden" bir "iz" olarak görülür. Bir iz, aralarında bir ilişki oluşması için diğer izlerle ilişkilendirilebilir. Bir iz bir şekilde etkinleştirilirse, onunla ilişkili diğer izler de etkinleştirilebilir. Bu olduğunda, bu izlerin temsil ettiği olaylar ya da deneyimler “akla gelir”, hatırlanır. Bununla birlikte, biraz farklı bir biçimde formüle edilen bu fikirlerin izi Descartes (1649, bkz. Descartes, 1961) ve Locke'un (Locke, 1690) yazılarında izlenebilir .
Bilginin kısa süreler için saklanması, aktivitede geçici bir artışla sağlanabiliyorsa , uzun süreli hafızanın beyinde yapısal değişiklikler gerektirdiğine inanılmaktadır. Descartes (1649; bkz. Descartes, 1961), izlerin "beyin gözenekleri olduğunu... [ki] diğerlerinden daha kolay yeniden açılma yeteneği kazanır ..." diye yazarken morfolojik değişim fikrini öngörmüştür . Freud (1895; bkz. Freud 1966) , nöronlar arasında engellerin varlığını öne sürdü . Nöronların aktivasyonunun, bu engellerin direncinde geri dönüşü olmayan bir azalmaya ve dolayısıyla bir hafıza izinin oluşmasına yol açtığına inanıyordu. Cajal'a (1911) göre kritik değişim yeni bağların oluşumunda yatarken, Tanzi (1893) ve Hebb (Hebb, 1949) iz oluştuğunda mevcut bağların güçlendiğine inanıyor . Daha yeni bir dizi kavram , bu temel görüşlerin varyantlarıdır (örneğin bkz . Minner 1957; Griffith 1966; Brindley 1967; Marr 1969).
Elbette, spekülatif kavramların egemen olduğu bu uzun dönemde, yine de teorik temellerde bazı değişiklikler oldu. Bazı yazarlar , kalıcı izin nöron devresindeki aktivitenin yankılanmasına bağlı olarak dinamik bir yapıya sahip olduğuna inanıyorlardı . Ancak birçok kişi ayak izini yapısal bir değişiklik olarak görüyor. Bu "yapısal" teoriler, yukarıda belirtilen fikirlere ek olarak şunları da önermektedir: beynin uydu hücrelerinde bir değişiklik, nöronların vücutlarındaki ribonükleik asidin (RNA) moleküler organizasyonunda ve spesifik lipoprotein komplekslerinde bir değişiklik nöronların zarlarında. Tabii ki, bu tam bir liste değil.
Bu görüşlerdeki farklılıklara rağmen, iki ortak noktaları vardır: nöronal değişiklikler davranıştaki değişiklikler tarafından değerlendirilir ve bu değişikliklerin doğrudan deneysel kanıtları en iyi ihtimalle göz ardı edilebilir ve en kötü ihtimalle tamamen yoktur (örneğin bakınız, Lashley, 1950) . . Bununla birlikte, çok sayıda teori ile deneysel verilerin azlığı arasındaki bu tür bir tutarsızlık, olağandışı bir şey değildir . "Yüksek zihinsel işlevlerin" altında yatan sinirsel süreçlere olan büyük ilgi, kaçınılmaz olarak bu süreçlerin doğası hakkında spekülatif tartışmalara yol açtı ve bu argümanlar gerçek sinir sistemini ilgilendirdiği için, doğal olarak, o zamanki bilgi durumuyla sınırlıydı. bir teorinin veya diğerinin geliştirilmesi . Deneysel verilerden ortaya çıkan derin anlayışın spekülatif akıl yürütmenin çok gerisinde kaldığını kabul ederken , 19. yüzyılın büyük nöroanatomistlerinin çalışmalarını gösteren Waldeyer'den bu yana 100 yıldan az bir süre geçtiğini hatırlamakta fayda var (bkz. Liddell, 1960). , aksonların terminal uzantılarının ilk olarak 1897'de Held tarafından açık bir şekilde tanımlandığı (Held, 1897) ve aynı yıl "sinaps" teriminin Sherrington tarafından bu uzantıların oluşturduğu bağlantıları belirtmek için icat edildiği şeklindeki "nöronal teori"yi kınadı. nöronların gövdeleri ile (Foster, 1897). O zamanlar, sinir sisteminin davranıştaki işleyişine ilişkin bilgi geliştirmenin en etkili stratejisi, çok çeşitli deneysel koşullar altında nispeten sabit kalan fizyolojik süreçleri incelemekti . Bu, bazı omurga reflekslerinin karakteristiğidir ve 1906'da Sherrington (Sherrington, 1906) parlak bir analizle bu reflekslerin omurilikte bütünleşme mekanizmalarını ortaya çıkardı . Öğrenme sürecinde ortaya çıkan davranış biçimleri bu özelliklere sahip değildir. Davranış öğrenme sürecinde değişir ve bu nedenle sinir sisteminin bir durumu olarak da düşünülmelidir. Beynin durumundaki bu değişikliğin doğasını anlamak için herhangi bir kararlı adım atılmadan önce , temel organizasyonunun derinlemesine incelenmesi gerekir. Yüzyılımızda sinir sistemi üzerine yapılan araştırmaların çoğu bu amaca yönelikti.
Bu açıdan bakıldığında, deneysel verilerin azlığı ile öğrenmenin sinirsel mekanizmaları ve bilgi depolama hakkındaki teorik fikirlerin zenginliği arasındaki çelişki daha anlaşılır hale geliyor.
Öğrenme ve hafızanın nöral temelleri üzerine yapılan araştırmalardaki gecikmenin bile bazı olumlu yönleri vardı. İlk olarak, son zamanlarda çok sayıda güçlü analitik yöntem geliştirilmiştir ve bunların sinir sistemi çalışmalarına uygulanması, onun yapısal ve işlevsel organizasyonuna ilişkin anlayışımızı kökten değiştirmiştir. İkincisi, davranışı tanımlama, analiz etme ve modellemede büyük ilerlemeler kaydedilmiştir. Tüm bu ilerlemeler, davranışın altında yatan süreçlerin derin ve ayrıntılı bir şekilde anlaşılmasına yol açmıştır . Çeşitli alanlarda, davranış üzerine çalışan araştırmacılar ile sinir sistemi üzerine çalışan araştırmacılar arasında ilgi alanları kesişmiştir. Bu birleşmenin sonuçlarından biri, artık beyinde bilgi kayıt biçimleri hakkındaki akıl yürütmenin deneysel verilere dayanmasıydı. Bu türden ilk veriler, bağımlılık araştırmalarında elde edildi.
hafıza ve alışkanlık
Bağımlılığın bazı davranışsal özellikleri
Humphrey (1930, 1933), Helix albolabris salyangozunun üzerinde hareket ettiği platform birkaç saniyede bir sallanırsa antenini geri çektiğini gösterdi. Yavaş yavaş, bu reaksiyon zayıflar ve sonunda koklea artık sallanmaya tepki vermez. Tepkinin bu kademeli olarak zayıflaması, alışkanlık adı verilen davranış değişikliğinin bir bileşenidir; bu fenomen hayvanlar aleminde yaygındır ( Humphrey, 1933; Thorpe, 1956). Alışmanın özellikleri , yönlendirme refleksi çalışmasında laboratuvar koşullarında kapsamlı bir şekilde incelenmiştir (Pavlov, 1927; Sokolov, 1960, 1965). Bu refleks, yeni bir uyarana tepki olarak ortaya çıkar ve uyaran tekrar tekrar sunulursa giderek zayıflar. Örneğin bir kedi aniden bir ses duyarsa sesin geldiği yöne bakar. Ses birkaç kez tekrarlanırsa , kedinin tepkisini uyandırmak için yavaş yavaş durur. Alışma , belirli sınırlar içinde, uyarana özgüdür, böylece tekrarlanan bir uyarana tepkinin kaybolmasından sonra, aynı etki başka bir uyaran tarafından üretilebilir. Bu uyaran bir süre sunulmazsa, tekrarlanan bir uyarana verilen yanıt eski haline getirilebilir .
hayvan için açıkça faydalı olan bir öğrenme sürecidir . Hayvan , başka sonuçları olmayan ve olmayabilecek tekrarlayan bir uyarana tepki vermeyi bırakır (Humphrey, 1933; Thorpe, 1956). Bununla birlikte, hayvan , tehlikeyi, yiyeceğin veya bir partnerin varlığını gösterebilecek yeni bir uyaran alma ve buna yanıt verme yeteneğini korur . Alışkanlığın altında yatan hafıza oldukça meraklıdır. Tepki vermeye gerek olmayan bir hatıradır; sönmüş reaksiyon geri yüklenirse, hayvanın * "unuttuğunu" söyleyebiliriz. Böyle bir hafızanın nöral analizi, davranıştaki alışmanın tezahürlerine paralel olarak , sinir fonksiyonlarının diğer bazı yönlerinde değişiklikler varsa, büyük ölçüde kolaylaştırılır. Bu tür değişikliklerin ilk kanıtı, beklenmedik bir kaynaktan, tamamlanmamış sinir sistemine sahip hayvanlar üzerinde yapılan bir dizi çalışmadan geldi.
fizyolojik analiz
Bir spinal köpek kaşınırsa, gıdıklanırsa veya omzun yakınındaki sırt derisine hafifçe vurulursa, uzuvun uyarı bölgesine doğru hareket ettiği bir dizi ritmik kaşıma hareketi olacaktır. Sherrington ( 1906) , derinin uzun süreli mekanik stimülasyonu ile tek refleksli kaşıma hareketlerinin genellikle yavaşladığını ve LC amplitüdünün azaldığını bulmuştur. Stimülasyon birkaç saniyeliğine durdurulursa, kaşınma refleksi geri yüklenir, ancak iyileşme her zaman bu kadar kısa bir aralıkta tamamlanmaz. Belirli sınırlar içinde, refleksin zayıflaması, cildin uyarılmış alanına göre spesifiktir: refleks kaybolduğunda, birkaç santimetre uzaktaki bir alanı uyararak kolayca indüklenebilir. Kaşıma refleksinin zayıflaması, bazı alışkanlık özelliklerine sahipti: reaksiyonun sönmesi , uyaranın uygulanma yerine nispeten özeldi; yok olma kas yorgunluğuna bağlı değildi ve muhtemelen deri reseptörlerindeki değişikliklere bağlı değildi; reaksiyon bir süre sonra düzeldi.
Sherrington'ın deneylerinde deri veya duyu sinirleri uyarıldı ve arka bacak hareketleri kaydedildi. Bu temelde, omurilikte meydana gelen süreçler hakkında sonuçlar çıkarıldı, ancak bu süreçler doğrudan incelenmedi. Alışkanlık sırasında nöral aktiviteyi kaydetme yeteneği birkaç yıl sonra ortaya çıktı ve önce omuriliğin değil beynin aktivitesi kaydedildi. İlk yaklaşımlardan biri, "uyaranın" tekrarlanan sunumu sırasında elektroensefalogram ritimlerindeki değişiklikleri incelemekti. İlk çalışmaların nesneleri insanlardı.
1 saniye
Resim: 1.1. Normal bir kedinin suprasylvian girusunun kortikogramı, 500 Hz'lik bir tona tipik bir bağımlılık uyanış tepkisi gösteriyor. yaklaşık 30 tekrardan sonra. A. 500 Hz'lik bir tonun ilk sunumuna tepki. Kaydın altındaki çizgi, uyaranın süresini gösterir; satırın sağında sunum zamanı saat, dakika ve saniyedir (10.45.00). Bir uyaranın sunumu üzerine, elektroensefalogramın aktivasyonu gözlenir. B. Uyaranın 36. ve 37. sunumlarında EEG. 38. denemede, bir önceki tonun (D) tekrarı eşliğinde 100 Hz'lik (V) yeni bir ton kullanıldı . E. 43. denemede 1000 Hz'lik yeni bir ton daha kullanıldı. Elektroensefalogramların sağ üst tarafında bulunan sayılar , her numunedeki aktivasyon reaksiyonunun süresini gösterir. (Sharpless, Jasper, 1956.)
(Berger, 1930; Knott ve Nepgu, 1941; Sokolov, 1960), anestezi uygulanmamış tavşanlar ( Ectors , 1936) ve kediler (Rheinberger ve Jasper, 1937). Bu değişiklikler Sharpless ve Jasper ( Sharpless ve Jasper, 1956) tarafından ayrıntılı olarak analiz edilmiştir. Bu deneylerde, anestezi uygulanmamış kedilerin uyumasına izin verildi ; uyku davranışsal ve elektroensefalografik kriterler ile değerlendirildi. Kediyi uyandırmak için 500 Hz'lik bir ton kullanıldı. Aynı zamanda, uyku elektroensefalogramının özelliği olan yavaş dalgaların yerini düşük voltajlı hızlı aktivite dönemleri aldı, yani elektroensefalogram etkinleştirildi (Şekil 1.1, L). Kedi tekrar uykuya daldığında ve uykunun elektroensefalogram özelliği geri gelir gelmez, ton tekrar açıldı. Tonun 20-30 kat tekrarlanmasıyla, uyaranlar kediyi uyandırmada giderek daha az etkili hale geldi ve elektroensefalogramın aktivasyonu gözlenmedi (Şekil 1.1, B). Hayvan 500 Hz'lik bir tonun tekrarında artık uyanmadığında, 100 veya 1000 Hz'lik bir frekansta başka bir ton kullanıldı (Şekil 1.1, C, E). Yeni tonların sunumu , hayvanın uyanmasına ve elektroensefalogramı etkinleştirmesine neden oldu . Tekrarlanan bir tona kortikal aktivasyon yanıtı alıştıktan sonra, yaklaşık 15 dakika veya daha uzun bir süre sonra aynı uyaran tarafından ortaya çıkarılabilir. Duraklamanın süresi kısmen önceki ton sunumlarının sayısına bağlıydı . Bazı durumlarda, sönmüş reaksiyonu tekrarlanan bir tona geri döndürmek için deneyler arasında birkaç günlük bir ara gerekliydi.
Elektroensefalogram değişiklikleri, uyarana özgü alışmanın birçok işaretiyle karakterize edilse de, Sharpless ve Jasper bu çalışmaların alışma sürecinin gerçek doğasını ortaya çıkarmadığına inanıyorlardı. Yine de bu çalışma en az iki açıdan önemliydi. Fizyolojik parametrelerdeki değişiklikler ile davranıştaki değişiklikler arasındaki ilişkiyi kurmuş ve bu süreci mikroelektrotlar kullanarak hücresel düzeyde analiz etmek için yöntemlerin geliştirildiği bir dönemde alışma sürecine dikkat çekmiştir .
Böyle bir analitik çalışma , tüm omurgalılarda önemli bir görsel alan olan optik çatıdaki tek nöronların aktivitesini kaydetti ( Hoth ve Hili, 1964–1966; Hoth, 1965; Masland ve diğerleri, 1971; Supader ve Bernan, 1972) . Memelilerde, retina çıkış nöronları -ganglion hücreleri- aksonları lateral genikülat gövdeye gönderir ve bu çekirdekteki hücreler görsel kortekse projekte olur. Ganglion hücrelerinin bir başka hedefi , iki yüksekliği olan görsel çatıdır - üstün kollikulus. Görsel çatının tek nöronları, görüntü retinanın karşılık gelen kısmına odaklanmışsa görsel uyaranlara yanıt verir . Retinanın bu alanı , üstün kollikulus nöronunun alıcı alanını oluşturur. Bazı hücreleri yalnızca görsel uyaranlara yanıt verir. Bununla birlikte, ismine rağmen görsel çatı diğer duyusal yollardan da girdi alır ( Honot ve Hili 1964, 1966; Jassik-Gershenfeld 1966; Gordon 1973; Drâger ve Hubel 1975).
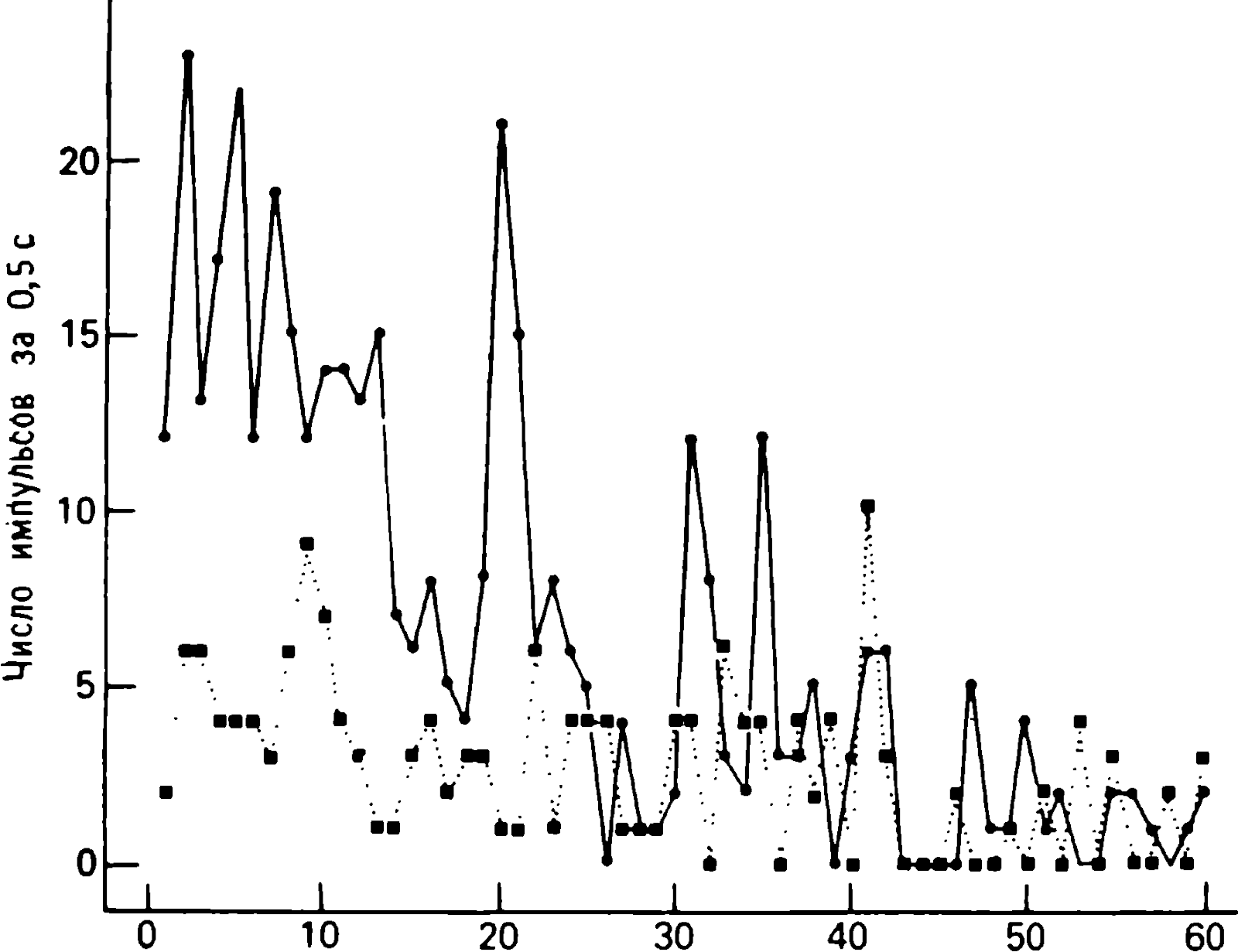
Uyuşturulmuş bir tavşanın optik çatısında kaydedilen bazı nöronların tepkileri , uyaran tekrarlandıkça zayıflar (Şekil 1.2). Tepkileri bu şekilde gösterilen hücre, herhangi bir özel duyusal uyarı olmadan kendiliğinden boşaldı ve omuz derisine yönlendirilen bir hava yaylımıyla bir uyarı yaylım ateşiyle karşılık verdi. Bu uyaran birkaç saniyede bir tekrarlandığında, tepki ilk 5-6 sunum için istikrarlı bir şekilde yeniden üretildi ve ardından zayıflamaya başladı. Yaklaşık 20 sunumdan sonra, uyaran artık spontan arka plan aktivitesinden farklı bir deşarja neden olmadı. Bu hücrelerin birçoğunda yanıt geri yüklenir.
2-140
25
Sıralı uyaran sunumları
Fis. 1.2. Anestezi uygulanmış tavşanın superior colliculus hücrelerinin reaksiyonları. Kafes, sağ omzu hedef alan bir hava yaylımına yanıt verdi. Patlama süresi 2 sn, uyaranlar arasındaki aralıklar 3 sn idi. Uyaranın her uygulanmasından önceki 0,5 saniyelik süre için spontan sivri uçların (kareler) sayısının ve ayrıca uyaranın uygulanmasından sonraki ilk 0,5 saniyedeki uyarılmış sivri uçların (daireler) sayısının uyaran dizisine bağımlılığı sayısı gösterilir. Tepki, sunulan uyaranların sayısı arttıkça kademeli olarak zayıflar. 23. sunumdan sonra, stimülasyon sırasında meydana gelen ani yükselme sayısı, stimülasyondan önce meydana gelen ani yükselme sayısından önemli ölçüde farklı değildir. (Ayaklar, Hili, 1966.)
Stimülasyon bir süre durursa infüze edilir. İyileşme yanıtının büyüklüğü, her zaman olmasa da genellikle iyileşme süresinin uzunluğuna bağlıydı; bilinen sınırlar dahilinde , ne kadar uzun olursa, iyileşme reaksiyonu o kadar güçlüydü. 1-2 dakikalık bir duraklama genellikle bir reaksiyonun meydana gelmesi için yeterli olmuyordu; bu , uyaranın önceki tekrarlarının sayısına bağlıydı .
Bununla birlikte, tepkinin zayıflaması yalnızca bir dereceye kadar uyarana özgüydü; kesinlik eksikliği, bazı uyaran genellemelerinin olduğunu gösterdi. Bu iki etki, ses uyaranlarına yanıt veren kollikulus hücresinin etkinliğiyle açıklanır (Şekil 1.3). Herhangi bir uyarı verilmeyen bir sürenin ardından 1000 Hz'lik bir ton uygulandı. Hücre bu tona ve 950 Hz'lik tona güçlü bir reaksiyonla tepki verdi (Şekil 1.3, L). Böyle bir testten sonra, hücreye 2 s aralıklarla 1000 Hz'lik bir ton sunuldu. Tepki kaybolduğunda 1000 Hz'lik bir ton yerine farklı bir frekanstaki tonlar sunuldu. 1000 Hz'lik bir tona yanıt
"TATLI PATATES"**
95°"
ıı.NHH_ _
w
1000 YIL
Yu00
1000XJ50 """"
Resim: 1.3. Uyaran genellemesine bir örnek. Tavşanın superior colliculus hücresi . Kayıtların altındaki çizgiler, belirtilen frekansın tonunun sunumunu gösterir. İlk başta, hücre 1000 ve 950 Hz (A) tonlarına güçlü tepki verir . 1000 Hz tona yanıt kaybolduğunda hücre 900 Hz tona yanıt verir (B, D) i. 1050 Hz'lik (D) bir tona "off^-tepkisi verir. Hücre 950 Hz tonuna (B) yanıt vermiyor . Bütün bu kayıtlar süreklidir. Stimülasyonun kesilmesinden birkaç dakika sonra hücre tekrar 1000 Hz'lik (E) bir tona yanıt verir. Tüm kayıtların düşük genlikli ani bir hücre gösterdiğine dikkat edin. Cevapları kaybolmuyor. Zaman kalibrasyonu - 0,5 sn. (Nogp'a göre, 1969.)
zayıf veya yok, ancak 900 Hz tonu bir reaksiyona neden oldu (Şekil 1.3, B, D) ve 1050 Hz tonuna bir dizi sivri uç eşlik etti (Şekil 1.3, E). Bu nedenle, sönme 1000 Hz tonuna özgüydü, ancak bu özgüllük ideal değildi : Başlangıçta bir yanıt da ortaya çıkaran 950 Hz tonu, 1000 Hz tonuna verilen yanıt söndükten sonra çalışmayı durdurdu. (Şekil 1.3, B). Başka bir deyişle, bazı genellemeler gözlemlendi . Birkaç dakikalık bir aradan sonra, hücre tekrarlanan ses tonuna tekrar yanıt verdi (Şekil 1.3, E).
2*

Sıralı uyaran sunumları
Resim: 1.4. Reaktivitenin kendiliğinden iyileşmesi. Tavşan optik çift koloni hücresi. Uyaran, her 2 saniyede bir sunulan 1000 Hz'lik kısa (0,5 saniye) bir tondur. 300 sn'lik bir duraklamadan sonra sunulan ilk uyaran serisi, uyarılmış reaksiyonlardır (sol eğri). 40 saniyelik bir duraklamadan sonra (ortadaki eğri) sunulan bir sonraki dizinin ilk birkaç uyaranıyla net tepkiler uyandırıldı , ancak daha sonra yanıtlarda yine hızlı bir zayıflama oldu. 20 saniyelik bir aradan sonra, hücre 20 uyarana güçlü bir şekilde yanıt verir ve ardından yanıtı azalır (sağdaki eğri). (Değil, 1970a.)
Davranışsal bir tepkinin yok olmasından sonra, genellikle kendiliğinden düzelir. Colliculus hücrelerinin tepkilerinin yok olmasından sonra bazen temelde benzer bir etki gözlemlenebilir (Şekil 1.4). Görünürde hiçbir sebep yokken, bu hücre birdenbire tekrarlanan bir uyarana şiddetli tepki vermeye başladı ve aynı anda tepki vermeyi bıraktı. Ayrıca bazen, beklenmedik bir dış uyaran sunarak, tekrarlanan bir uyarana davranışsal tepkiyi geri yükleyebilirsiniz (Humphrey, 1933; Lehner, 1941). Bu etki en çok böyle bir uyaran sabit bir uyaranın birbirini izleyen sunumları arasına eklendiğinde belirginleşir . Anestezi uygulanmış tavşanda optik kollikulusun nöronlarının deşarjları kaydedilirken bu tür bir reaktivitenin geri kazanılması gözlenmedi . Ancak bu etki, çekirge Schistocerca gregaria Forskal'ın beynindeki nöronal deşarjları kaydederken gözlendi (Horn ve Rowell, 1968; Rowell ve Horn, 1968). Kayıt, beynin tritocerebral lobunun nöronlarından yapıldı . Gösterilen nöronun karakteristik bir özelliği, tekrarlanan stimülasyon üzerine görsel alan boyunca hareket eden siyah bir diske verilen yanıtta hızlı bir düşüş oldu (Şekil 1.5). Çekirge beyni , servikal bağ adı verilen iki sinir lifi demeti ile ventral sinir kordonuna bağlanır. Hareket eden diske verilen yanıt zayıfladığında, kontralateral servikal bağa kısa bir elektriksel uyarı patlaması uygulandı . Tekrarlayan bir görsel uyarana tepki önemli ölçüde arttı (Şekil 1.5). İpsilateral bağın, yani deşarj elektrodunun bulunduğu taraftaki bağın uyarılmasının, görsel uyaranla uyarılan tritocerebrum hücresinin reaksiyonları üzerinde hiçbir etkisi olmamıştır. Kontralateral bağın uyarılmasının güçlendirici etkisi kademeli olarak azaldı (Şekil 1.5, eğriler IV ve V). Benzer değişiklikler, davranışsal tepkilerin dış uyaranlar tarafından tekrar tekrar engellenmesi üzerine gözlenir (Lehner, 1941).
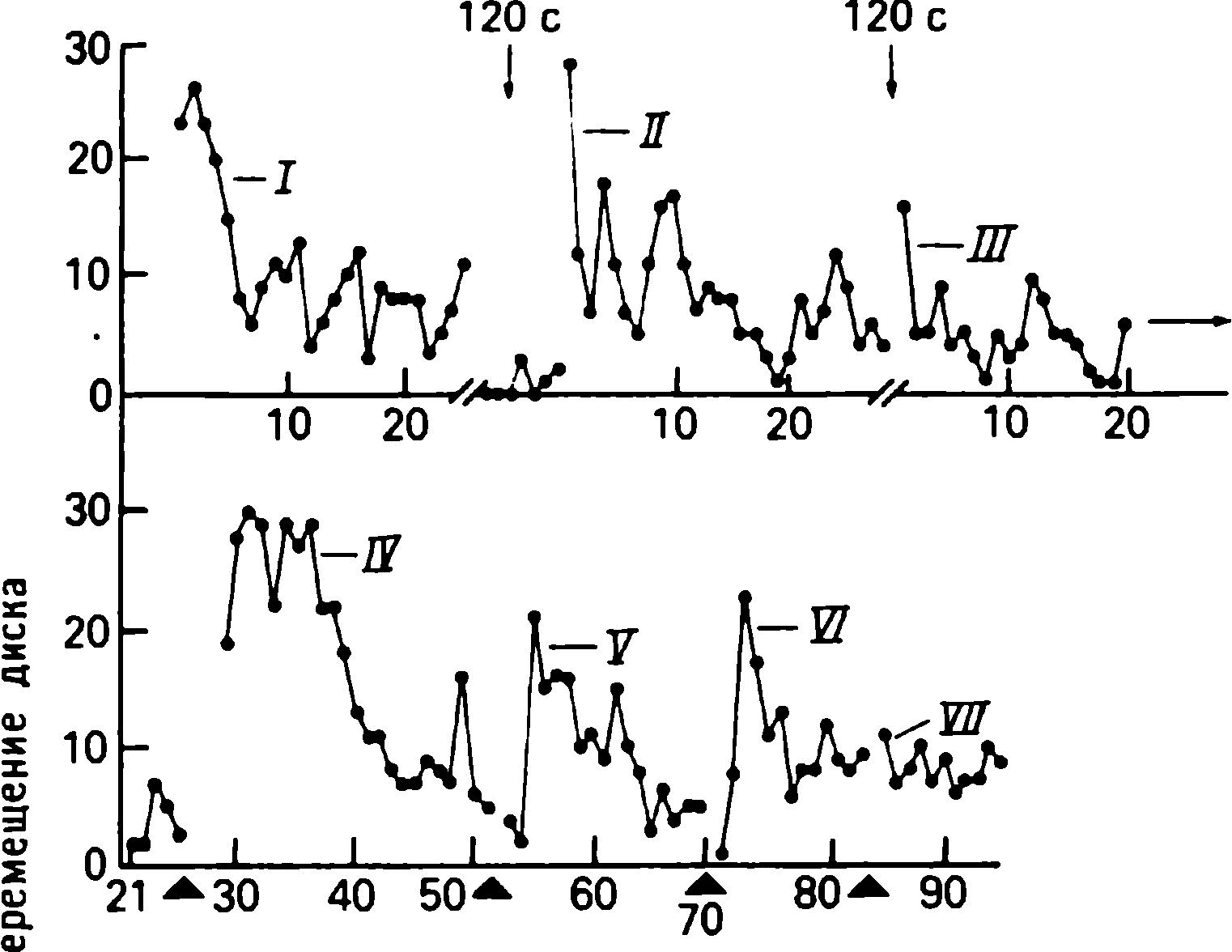
Tritocerebral hücrenin siyah diskin hareketine verdiği yanıt kaybolduğunda, genellikle stimülasyonun bir süre durdurulmasıyla eski haline dönebilirdi. Ancak, böyle bir iyileşme her zaman gözlenmedi. Örneğin, reaksiyonları Şekil 1'de gösterilen bir hücre. 1.6, siyah diskin alıcı alanı boyunca 10 saniyelik aralıklarla yukarı doğru hareketine yanıt verdi. 23 dakikalık bir aradan sonra, hücre uyarana güçlü bir şekilde tepki verdi ve ardından tepki azaldı. İki uyaran dizisi arasındaki 5 saatlik bir aralıkta, iyileşme çok hafifti (Şekil 1.6, sırasıyla eğriler I ve II ). Böylesine uzun süreli bir depresyonun, deney hayvanının durumundaki genel bir bozulmaya bağlı olup olmadığını belirlemek gerekliydi. Böyle bir açıklama olası değildir, çünkü hücre, ikinci dizi yukarı hareketlerden hemen sonra sunulan diskin ileri hareketine (Şekil 1.6, beyaz üçgen) ilk uyaran dizisinin sunumundan önceki kadar güçlü yanıt vermeye devam ettiğinden (Şek. 1.6, siyah üçgen).
1960'larda, hipokampus (Vinogradova, 1970) ve beyin sapı (Lettvin ve diğerleri, 1961; Grüsser-Cornehis ve diğerleri, 1963; Bell ve diğerleri, 1964) ve omurilik (Buchwald ve diğerleri, 1965; Spencer ve diğerleri, 1966a, b, c). Bazı omurgasızlarda beyin nöronlarının ayrıntılı çalışmaları da yapılmıştır (Holmgren ve Frenk, 1961; Roberts, 1962; Hughes ve Tais, 1963; Horridge ve diğerleri, 1965; Bruner ve Tauc, 1966; Krasne ve Roberts, 1967). ug gelişimi
- 15 dakika

Sıralı uyaran sunumları
Resim: 1.5. Bir uyaranın tekrarı üzerine çekirgelerde azalan reaksiyonlar ve ipsilateral değil, kontralateral servikal bağın uyarılmasından sonra bunların düzelmesi. Görsel uyaran, 1.4 saniyede 40° hareket ettirilen bir diskti. Ardışık hareketler arasındaki aralık 10 sn idi. Apsis ekseninin altındaki siyah ve beyaz üçgenler , ardışık disk sunumları arasında sırasıyla kontralateral ve ipsilateral bağlara uygulanan darbe patlamalarını gösterir. Okların üzerindeki sayılar, birbirini izleyen uyaran grupları arasındaki zaman aralıklarını gösterir. X eksenine paralel oklar , stimülasyonun kesintisiz devam ettiğini gösterir. Eğri I ve II arasındaki eğri , stimülasyon yokluğunda deşarjların kontrol sayısıdır. (Rowell r Noth, 1968.)

Sıralı uyaran sunumları
Resim: 1.6. Tepkilerde sürekli uyarana özgü azalma. Her daire, diskin 10 s aralıklarla yukarı doğru hareketinin neden olduğu sivri uçların sayısını temsil eder . İlk hareket serisinden (eğri /), hiçbir stimülasyonun uygulanmadığı 23 dakikalık bir duraklama geldi. 1. ve 2. hareket serisi arasındaki aralık 5 saattir II . eğrideki ilk uyaran, / (71 sivri) eğrideki ilk boşalmaya kıyasla yalnızca nispeten zayıf bir boşalmaya (25 yükselme) neden olur . 2. yukarı vuruş serisinden sonra ilerlemek, neredeyse ilk yukarı hareket serisinden (siyah üçgen) önce olduğu kadar çok sivri uçlara (beyaz üçgen) neden olur. (Rowell, Noth, 1968.)
Hem nöronal hem de davranışsal seviyelerde ekstra uyaranlara yanıt olarak iyileşmenin özgüllük derecesi , kapsamı ve özellikleri zaman içinde önemli ölçüde değişir (Hinde, 1970), ancak sürecin bazı ortak özellikleri birçok farklı hayvan türünde kendini gösterir. Dahası, nöronal değişikliklerin doğası birçok açıdan davranıştaki alışmaya benzer. Nöronlardaki değişiklikler ile davranışsal tepkilerdeki değişiklikler arasında olası bir nedensellik ilişkisi sorusunu ele almadan önce, nöronlarda meydana gelen değişikliklerin mekanizmaları üzerinde durmak gerekir.
Hücresel mekanizmalar
Sherrington ( 1906) bir çalışmasında kaşıma refleksindeki zayıflamanın kas yorgunluğuna bağlı olmadığını açıkça gösteren verileri aktarmaktadır,
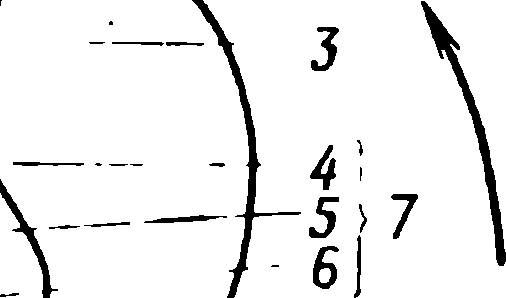
Resim: 1.7. Multisinaptik refleks yayının bazı bileşenlerini gösteren memeli omuriliğinin enine kesit diyagramı . Deri stimülasyonunun neden olduğu afferent impulslar , afferent lifler tarafından omuriliğe taşınır (2). Bu liflerin uçlarından bazıları ara nöronlarla sinaptik temaslar oluşturur (3). Omuriliğin gri maddesinde (1) yer alan bu nöronlar impulsları sadece motor nöronlara (8) iletir. Motor nöronların aksonları (9) omurilikten çıkar ve fleksör kasları innerve eder. Omurgalı bir nöronun (7) ana parçaları soma veya perikaryon olarak da adlandırılan hücre gövdesi ( 4), akson (5) ve biri (6) şematik olarak gösterilen çok sayıda sinaptik bağlantıdır . Bu nöronun dendritleri gösterilmemiştir. Dendritler , genellikle geniş dallar oluşturan ve omurgalılarda genellikle somadan ayrılan sitoplazmik süreçlerdir . Diyagram ölçekli çizilmemiştir: perikaryanın çapları 10 ila 70 µm arasında değişirken, omuriliğin milimetre cinsinden ölçülen ortalama bir çapı vardır; gerçek boyutları farklı türlerde büyük farklılıklar gösterir.
bu reflekse neden olan Bu kaslardan birini innerve eden motor sinirin doğrudan uyarılması üzerine iyi kasıldı , ancak refleks kayboldu. Sherrington, kasları innerve eden motor nöronların pratik olarak yorulmaz olduğuna inanıyordu, ancak bu motor nöronların reseptörler ve afferent yollar tarafından gerçekleştirilen kontrolünün daha az güçlü hale geldiğine inanıyordu; uzun süreli uyarılma ile motor nöronlar üzerindeki etkileri zayıflar. Sherrington'a göre yorgunluğun gelişme alanı, deride ve cildi omuriliğe bağlayan duyu sinirlerinde değil, motor nöronların gövdelerinin bulunduğu omurilikte bulunur. Yorgunluğun sinaps bölgesinde geliştiğine inanıyordu .
1898'de Sherrington (Sherrington, 1898), fleksiyon refleksinin genliğinin, buna neden olan uyaranın tekrar tekrar sunulmasıyla da azaldığını gözlemledi. Daha sonra Prosser ve Hunter (Prosser, Hunter, 1936) bu refleksin gücündeki değişimi ayrıntılı olarak incelediler. Refleks, karşılık gelen duyusal sinirin derisinin veya liflerinin uyarılmasıyla indüklendi (Şekil 1.7). Bu tür bir uyarım , belirli kasların - uzvun fleksörlerinin - kasılmasına yol açar . Kasılmalar doğrudan omuriliğin motor nöronları tarafından kontrol edilir. Spencer ve diğerleri (1966a, b) ve Wickelgren (1967a) , kedilerde , bir uyaranın tekrar tekrar sunulması üzerine bu refleksin genliğinin nasıl azaldığını incelediler. Omurilikte afferent impulsların dağılımında bir değişiklik buldular, ancak bu değişiklik motor nöron yanıtlarındaki azalmayı açıklamak için yeterli değildi. Sherrington'ın, motor nöronların kontrolünün zayıfladığı, ancak kendilerinin - "nihai ortak yol" - görece yorulamaz olduğu görüşü, Spencer ve arkadaşlarının deneylerinde doğrulandı (Spencer ve diğerleri, 1966c). Bu araştırmacılar , mikroelektrotları doğrudan omuriliğin motor nöronlarına enjekte ettiler. Azalan yanıtların, nöronları etkileyen azalmış uyarılma ile ilişkili olduğunu bulmuşlardır ; bu zayıflama, motor nöronun giriş sinyaline yanıt verme yeteneğindeki herhangi bir değişikliğe bağlı değildi. Bu nedenle, omuriliğe afferent girdi ile buradan efferent çıktı arasında bir yerde bir iletim bozukluğu gelişmelidir (Wickelgren, 1967b; Wall, 1970).
İletim bastırma çeşitli şekillerde meydana gelebilir . Olası yollardan biri, frenleme süreçlerine bağımlılıktır. Örneğin, aksiyon potansiyeli patlamaları bir nöron zincirinden geçtiğinde , devrenin inhibitör dalında aktivitede bir artış meydana gelebilir. Bu kademeli yavaşlama, karşılıklı bir şekilde hareket edebilir ve kademeli olarak devredeki iletimi bloke edebilir. Böyle bir mekanizma bazı sistemlerde kesinlikle mümkündür, ancak neredeyse evrensel değildir. Bruner ve Tock tarafından özellikle önemli bir dizi deney gerçekleştirildi (Burner ve Thais, 1966). Deniz yumuşakçası Ariusia'nın sol plevral ganglionunun dev bir nöronundan gelen potansiyellerin hücre içi kaydını gerçekleştirdiler . Başın sınırlı bir bölgesine bir damla su düştüğünde, dokunaçlardan biri veya her ikisi geri çekildi. Yaklaşık 10 sn aralıklarla su damlalarının etkisi altında, bu reaksiyon giderek azaldı . Stimülasyon bir süre durdurulursa, reaksiyon bir dereceye kadar eski haline döner ve bu iyileşme ek faktörlere bağlıdır. Tepki, uyaranların sunumunda herhangi bir kesinti olmadan iyileşebilir; bunun için yumuşakçanın başını gıdıklamak veya siniri uyarmak yeterliydi. Bir su damlasının dokunaçların kasılmasına neden olduğu anda, dev hücrede yüksek amplitüdlü eksitatör postsinaptik potansiyel (EPSP) kaydedildi (Şekil 1.8, L). EPSP'nin genliği, dokunaçların kasılma büyüklüğündeki değişikliklere paralel olarak değişti. Kaydedilen EPSP karmaşıktı ve büyük olasılıkla birkaç presinaptik nörondan gelen birkaç üniter EPSP'den oluşuyordu. Bununla birlikte, üniter EPSP'deki düşüşün niteliği
Uyaran sunum sırası

ZOOms
AB _
Resim: 1.8. Aplysia'da tekrarlanan stimülasyonun EPSP genliği üzerindeki etkisi. A. Sol ön dokunaç I'in yakınındaki bir yumuşakçanın kafasına 10 saniyelik aralıklarla düşen su damlalarıyla ortaya çıkan karmaşık bir EPSP'nin kayıtları — 15 uyaranlık ilk seri; II - on dakikalık bir aradan sonra EPSP'nin iyileşmesini gösteren ikinci seri; III - stimülasyon sıklığını değiştirmeden, ancak sol arka pedal sinirinin 2 s (1 s'de 5 stimulus) ritmik stimülasyonundan sonra ikinci serinin devamı. EP amplitüdünün geri kazanılmasına dikkat edin 5. Stimülasyon etkilerinin üniter bir EPSP'nin iyileşme süreci üzerindeki etkisi. EPSP, sol plevrovisseral konjonktivanın pupa liflerinin 10 sn aralıklarla uyarılmasıyla indüklendi. Y ekseni, EP genliğini başlangıç değerinin yüzdesi* olarak gösterir; apsis ekseni boyunca, saniye cinsinden süre
(Şekil 1.8, B) , karmaşık EPSP ile aynıydı. Üniter EPSP'lerin tek bir sinaptik temastan önlendiği varsayılmıştır. Bu durumda üniter potansiyel, presinaptik sinir lifinde tek bir impulsun deşarjına bağlıydı. Bruner ve Tok, böyle bir EPSP'deki düşüşün, postsinaptik hücrenin durumundaki bir değişiklikten bağımsız olduğuna dair güvenilir kanıtlar sundular; bu veriler , sinaptik iletim bozukluğunun presinaptik mekanizmasına tanıklık etti.
Akson boyunca terminale ulaşan kimyasal sinapslardaki sinir impulsu, postsinaptik hücrenin uyarılabilirliğini artıran veya azaltan bir aracıyı serbest bırakır. Uyarıcı etkinin kanıtı , postsinaptik hücrenin depolarizasyonudur. Depolarizasyon kendini EPSP şeklinde gösterir. Depolarizasyon miktarı yeterliyse, akson boyunca postsinaptik nörona yayılan bir aksiyon potansiyeli oluşabilir . İnhibisyon, "postsinaptik hücrenin zar potansiyelinin ters yönde kayması" şeklinde kendini gösterir. Bu inhibitör postsinaptik potansiyeldir (IPSP). Aracı, muhtemelen sinapslarda bulunan küçük keseciklerde paketlenmiş moleküler kompleksler biçiminde salınır (incelemeler için bkz. Katz, 1969; Kelly ve diğerleri, 1979). Bruner ve Tok, gözlemledikleri yanıtın zayıflamasının, nörotransmitter salınımındaki azalma nedeniyle sinaptik iletimdeki bir depresyonun sonucu olduğunu öne sürdüler . Hipotezlerine göre bu düşüş, terminallerdeki nörotransmiterlerin tükenmesinden ve onun yavaş mobilizasyonundan kaynaklanıyordu. Sinapsta ortaya çıkan iletim blokajı, sinir sisteminde bilgi iletiminin aktiviteye bağlı veya otomatik olarak oluşturulan bir depresyonunun bir örneğidir (Horn, 1967). Böylece, bir nöronun aktivitesi, kendi sinyal iletme yeteneğini engeller.
onlar tarafından gözlemlenen üniter EPSP'lerdeki değişikliklerin, ek bir nöron zincirinin katılımı olmadan yalnızca presinaptik liflerdeki değişiklikleri yansıttığı varsayımına dayanmaktadır . Ancak Bruner ve Tok , notlarının tek bir kişiden alındığı iddiasında ihtiyatlıydılar . Böyle oldukça önemli bir temas , kalamar Loligo vulgaris'in (Young, 1939) yıldız ganglionunda bulunur. Bu ganglionda internöronlar yoktur Ek bir avantajı da, pre- ve postsinaptik elemanların boyutları çok büyük olduğundan, impuls iletimi ile ilişkili potansiyel değişikliklerin de büyük olması ve hücre dışı elektrotların uygun şekilde yerleştirilmesiyle tespit edilebilmesidir (Bullock , 1948). Ek olarak, bu sinaps yoluyla iletim iyi çalışılmıştır.
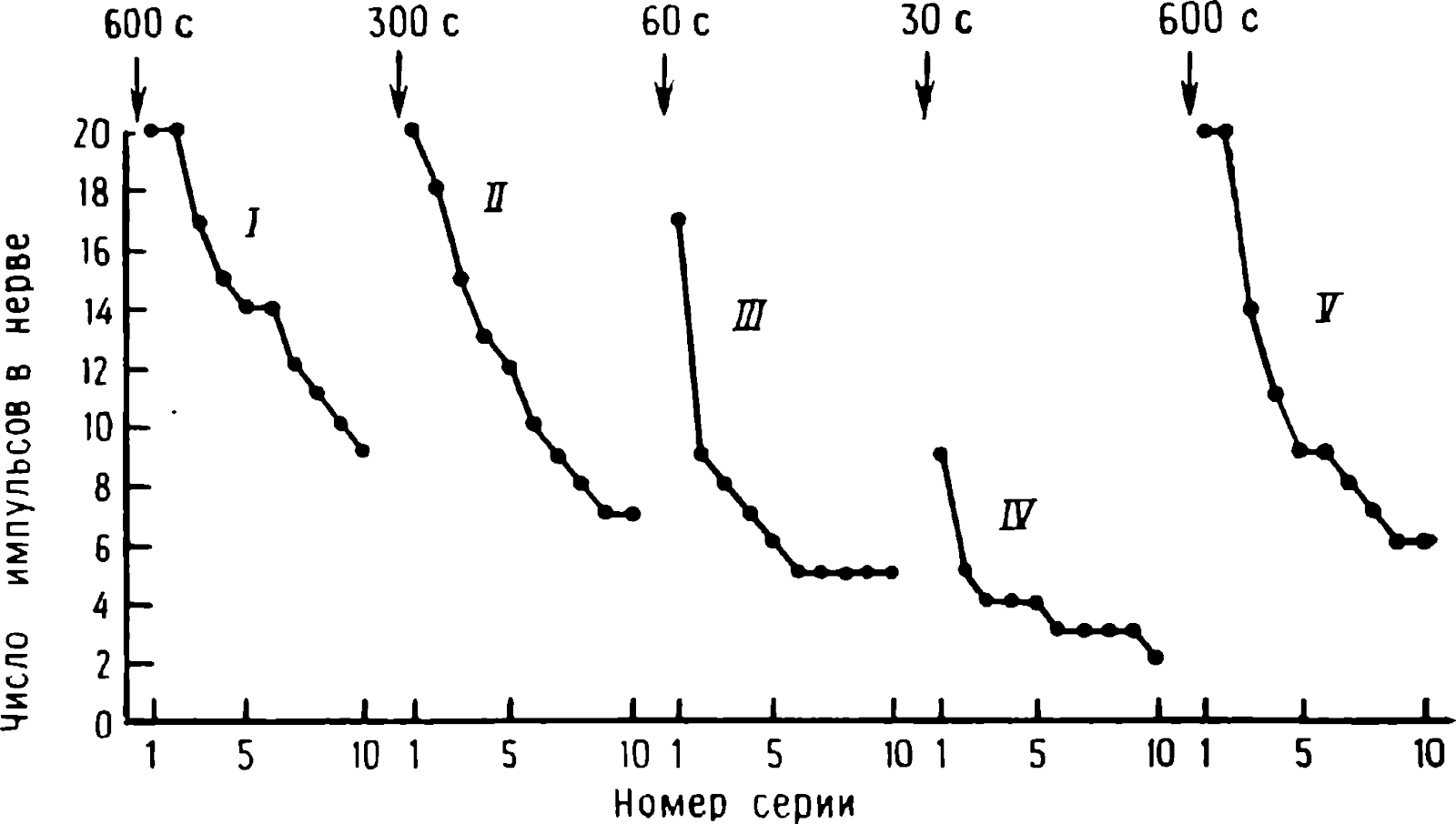
Resim: 1.9. Farklı iyileşme sürelerinin yıldız gangliyonunda iletim üzerindeki etkisi. Her uyaran dizisi, 0,5 s için 40 darbe/s frekansında iletilen 20 elektriksel darbeden oluşuyordu . Ardışık seriler arasındaki aralık 10 sn idi. Okların üzerindeki rakamlar iyileşme süresinin süresini gösterir. (Horn ve Wright, 1970.)
Presinaptik lifin ritmik uyarımı üzerine dev sinapsta iletim durur mu ? Eğer öyleyse, tepkinin azalmasının ve iyileşmesinin doğası Şekil 1'de gösterilene benzer mi? 1.5 (eğriler I-III) ve şek. 1.8, B? Bu soruları cevaplamak için yıldız ganglion eksize edildi (Horn ve Wright, 1970). Presinaptik lifleri içeren prednerv, elektriksel impulslarla uyarıldı ve dev postsinaptik aksondaki yanıtlar, hücre dışı elektrotlarla kaydedildi. Ön sinir , her 10 saniyede bir tekrarlanan kısa uyarı dizileriyle uyarıldığında, her bir dürtü, postsinaptik aksonda bir yükselmeye neden oldu ve iletim neredeyse sonsuza kadar devam etti. Bir serideki dürtü sayısındaki 20'ye kadar bir artışla, 0,5 s'de ilk serinin her bir darbesi, postsinaptik hücrede bir artışa neden oldu (Şekil 1.9, eğri /). Bununla birlikte, yanıt zayıfladı ve ön sinire uygulanan 20 uyarılık beşinci seri, postsinaptik lifte yalnızca 14 sivri uç oluşmasına neden oldu. Stimülasyonun kesilmesinden sonra yanıt eski haline döndü ve iyileşme derecesi farklıydı. Belirli sayıda uyaran serisi için, toparlanma miktarı duraklamanın süresine bağlıydı. Seri tekrarlandığında, Aplysia'da gözlenen duruma benzeyen (Şekil 1.8, B) stimülasyonun etkileri birikmiştir (Şekil 1.9). Yıldız ganglionun prenervesinin postsinaptik sinirde sivri uçlar oluşturmak için elektriksel stimülasyonunun verimsizliğindeki bir artış , EPSP değerinde ilerleyici bir düşüşle ilişkilendirildi.
EPSP değerindeki bu değişikliklere neden olan faktörler birçok araştırmacı tarafından incelenmiştir. Aynı zamanda pre- ve postsinaptik temas elemanlarına mikroelektrotlar yerleştirildi ve pre-sinire uyarılar uygulandı. Bu koşullar altında: postsinaptik tepkinin genliği azaldı (Bullock ve Hagiwara, 1957; Bryant, 1959), ancak aksiyon potansiyelleri presinaptik terminal boyunca yayılmaya devam etti . Böylece tekrarlanan stimülasyon, presinaptik terminale gelen aksiyon potansiyeli ile EPSP'lerin oluşumunda ifade edilen nörotransmiterin salınması arasındaki bağlantının bozulmasına yol açar. Kalamar dev sinapsında bu potansiyelin presinaptik terminalin durumuna bağlı olduğuna dair iyi belgelenmiş* kanıtlar vardır . Bu, iletimin bozulmasının tamamen veya esas olarak temasın presinaptik kısmında gerçekleştiğini düşündürür (Katz ve Miledi, 1969). Bu aktarımın bozulması için çeşitli olası moleküler mekanizmalar düşünüldü ve bazıları reddedildi (Horn ve Wright, 1970). Sinaptik terminalde bir aksiyon potansiyeli ortaya çıktığında kalsiyum iyonlarının buraya girdiği bilinmektedir . Bu süreç, nörotransmitter salınımı için gereklidir (Katz ve Miledi, 1967; Miledi, 1973), bu nedenle, kalsiyum iyonlarının konsantrasyonu ve hareketindeki değişiklikler, ön sinirin tekrar tekrar uyarılmasıyla iletimdeki değişikliklerin olası bir açıklaması olarak kabul edilmiştir (Horn ve Wright, 1970; Horn, 1970b).
Bruner ve Tok, inceledikleri dev hücrenin dokunaç retraksiyonunun refleks yayına girmediğini fark ettiler. Ancak bu refleksin genliğindeki değişimin kinetiği EPSP'nin kinetiğine benzerdi. Bu benzerliğe dayanarak, sinapslardaki değişikliklerin davranışta alışmaya neden olan temel mekanizmalardan biri olabileceğini öne sürdüler . Sinapslardaki ve davranıştaki değişiklikler arasındaki bağlantı, bu davranışsal yanıtı doğrudan düzenleyen nöron zincirindeki iletimi incelemek mümkün olduğunda daha net hale gelecektir.
refleks uzuv fleksiyonunu düzenleyen nöronların ana zinciri bilinmektedir. Wickelgren (Wickelgren, 1967a, b) ve Wall (Wall, 1970) , görünüşe göre, refleks yayının belirli bir bağlantısında meydana gelen refleksin sönmesiyle ilişkili iletim ihlalinin olduğunu gösteren veriler elde ettiler. Bununla birlikte, omurilik sinir ağlarının aşırı karmaşıklığı nedeniyle iletim bozukluklarına dahil olan hücresel mekanizmaların incelenmesi zordur. Bu arada Kupferman ve Kandel (1969) , Aplysia'da solungaç geri çekme refleksinin uygulanmasında yer alan nöral devrenin anatomisini aydınlattı . Vücudun yüzeyine - mantonun kenarı veya sifon - dokunulduğunda solungaçlar, manto ve sifon kasılır. Uyaranın tekrar tekrar sunulması üzerine tepki zayıflar. Stimülasyonun geçici olarak kesilmesiyle veya fazladan bir uyaranın sunulmasıyla (örneğin, refleksin alıcı alanının bir kısmına elektrik şoku) geri yüklenir (Pinsker ve diğerleri, 1970). Solungaçların refleks geri çekilmesinin zayıflaması, duyu nöronlarının afferent deşarjlarındaki değişikliklere bağlı değildi; solungaçları kontrol eden motor nöronun doğrudan elektriksel uyarımı sürekli olarak güçlü bir solungaç kasılması tepkisi ortaya çıkardı (Kupfermann ve diğerleri, 1970). İletimin bozulmasının refleks arkın duyusal ve motor nöronları arasında meydana geldiği bulunmuştur . Davranışsal yanıttaki bir azalma , motor nörondan EPSP çıkışının genliğindeki bir azalma ile ilişkilendirilmiştir (Castellucci ve diğerleri, 1970). Temas muhtemelen monosinaptik olduğundan, bu yazarlar EPSP'deki düşüşü , yaklaşık olarak Bruner ve Tok'un önerdiği şekilde (Bruner ve Tais, 1966) duyusal nöronun terminallerinden aracı çıkışındaki bir azalmayla açıkladılar.
Klein ve diğerleri ( 1980) , bir duyu nöronunun terminallerindeki verici salınımı ile iyonik akımlar arasındaki ilişkiyi araştırmaya karar verdiler . EPSP'deki ilerleyici düşüşün, gelen kalsiyum akımındaki bir azalmaya eşlik ettiği gösterilmiştir . Dinlendikten sonra iletimin restorasyonu, bu akımdaki bir artışla ilişkilendirildi. Hücre gövdesindeki kalsiyum akımındaki değişiklikler, dxon terminallerinde meydana gelen değişiklikleri doğru bir şekilde yansıtıyorsa, o zaman bu sonuçlar, alışmayla ilgili sinaptik depresyonun kalsiyum iyonlarının hareketinin bir sonucu olduğu şeklindeki önceki öneriyi (s. 29) destekler.
Alışkanlığı inceleyen birçok çalışmada dikkatler , tepkisellikteki kısa vadeli değişimlerin analizine, yani birkaç dakika veya saat sonra iyileşme meydana geldiğindeki değişimlere odaklanmıştır. Bununla birlikte, birkaç gün veya hafta süren çok sayıda alışkanlık örneği vardır (örneğin bkz. Hinde, 1960; Glaser ve Whittow, 1957; -Griffin ve Pearson, 1967). Yukarıda anlatılana benzer bir nöral mekanizma aynı zamanda uzun vadeli bir alışkanlık da sağlayabilir mi? Locust tritocerebrum'da nöronların tepkisinde 5 saate kadar süren uyarana özgü bir azalma gözlendi (Şekil 1.6), ancak daha uzun süreli nöronal değişiklikler de bulundu. Castellucci ve diğerleri ( 1978), Aplysia'daki solungaç retraksiyon refleksinin, uygun stimülasyona alıştıktan sonra birkaç hafta boyunca azalmış kalabileceğini gösterdi . Refleksin alıcı alanında aynı yere art arda beş veya daha fazla gün boyunca tekrarlanan uyaranların uygulanmasından ibaretti. Bu prosedürden sonra, duyusal ve motor nöronlar arasındaki iletim üç haftaya kadar azalmış olarak kaldı.
Ekstra tahriş edici, genellikle güçlü ve hatta acı verici , sönmüş bir davranışsal tepkiyi geri getirebilir. Yalnızca söndürülmüş reaksiyon geri yüklenirse, bu etkiye disinhibisyon denir. Bununla birlikte, kural olarak, diğer reaksiyonların gücü de aynı anda artar. Artan reaktivitenin bu daha genel etkisine genellikle "duyarlılık " denir (Thompson ve Spencer, 1966). Bazı bağımlılık biçimlerinin altında sinaptik depresyon yatıyorsa, ekstra bir uyaran iletimi nasıl geri getirir?
Sinapsta salınan aracı miktarı, resinaptik terminalin polarizasyon seviyesine bağlı olarak değişebilir. Örneğin, kalamar dev sinapsındaki sinaptik potansiyelin genliği, ön sinir sürekli olarak kısa elektriksel impulslarla uyarılırsa azalır . EPSP genliği, presinaptik terminalin dinlenme potansiyelini artırarak veya hiperpolarize ederek geri yüklenebilir ( Hagiwara ve Tasaki , 1958; Takeuchi ve Takeuchi, 1962; Miledi r Slater, 1966). Bu etkinin salınabilen aracı madde miktarındaki artıştan kaynaklandığına inanılmaktadır. Kalamarın yıldız ganglionu hakkında elde edilen verilere dayanarak, ekstra bir uyaranın uygulanmasından sonra reaksiyonların restorasyonunun , depresyon halindeki presinaptik terminalin hiperpolarizasyonundan kaynaklanabileceği öne sürülmüştür (Hot, 1967). ). Güçlü veya zararlı bir uyaran , beyin sapı retiküler oluşumu olarak bilinen oldukça dallanmış bir sistemi harekete geçiriyor gibi görünmektedir (Hobson ve Scheibel, 1980). Bu spesifik olmayan sistem, büyük olasılıkla, tüm omurgalılarda mevcuttur; benzer sistemler omurgasızlarda da bulunabilir (Rowell, 1970). Böyle bir sistemin nöronları, depresyon halindeki terminalleri hiperpolarize ederse, ekstra bir uyaranla aktivasyonu iletimi yeniden sağlayabilir (Şekil 1.10). Duyusal uyaranlardan bağımsız olarak spesifik olmayan bir sistemin aktivitesinde ani bir artış aynı etkiye sahip olabilir.Bu durumda, tekrarlanan bir uyarana verilen tükenmiş tepkinin "kendiliğinden" bir restorasyonu olacaktır. Hem nöronal (Şekil 1.4) hem de davranışsal seviyelerde benzer iyileşme bulunmuştur . Spesifik olmayan bir sistem tarafından oluşturulan bağlantılar , mutlaka nöronlar arasındaki doğrudan sinaptik temaslar yoluyla gerçekleştirilmez. Nöromodülatörler olarak adlandırılan bazı maddeler spesifik olarak salınmayabilir.
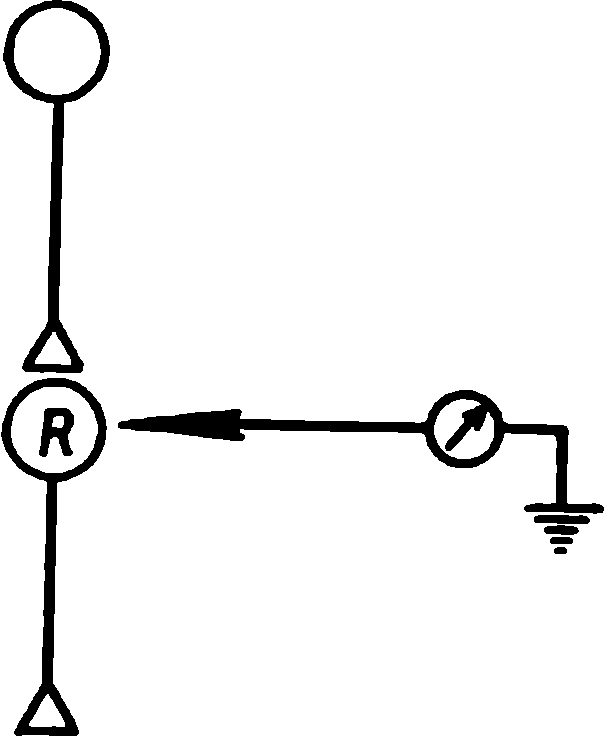
Resim: 1.10. Disinhibisyon veya sensitizasyonun olası nöral mekanizması . Uyaran, P hücresinde bir dizi deşarja neden olur . Bu sinyal , aktivitesi bir mikro elektrot tarafından kaydedilen R hücresine iletilir . Uyaran tekrarlandığında P hücresi boşalmaya devam eder, ancak R hücresinin tepkisi azalır. İletim blokajının , P hücre terminallerindeki sinaptik depresyondan kaynaklandığı varsayılmaktadır.Güçlü bir ekstra uyaran , iletimi geri yükler . Bu modelde, bu etkiye kök retiküler oluşumu tarafından aktive edilen inhibitör bir nöron aracılık eder. Beyaz üçgenler azalmayan uyarıcı sinapsları belirtir ; gölgeli - azaltıcı uyarıcı sinaps; siyah — inhibe edici azalmayan sinaps. (Değil, 1967.)
fiziksel sinaptik temaslar ve bunların yakınında ve bir dizi sinapsta iletimin doğasını değiştirir (Shepherd, 1979). Dudel (1965) tarafından gösterildiği gibi , indolamin serotonin, kanserin nöromüsküler kavşağında aracı salınımını arttırır. Aplysia'da karşılık gelen ekstra uyaran, depresyon halindeki sinapslardaki davranışsal yanıtı ve iletimi geri yükler (Burner ve Tais, 1966; Pinsker ve diğerleri, 1970). Hassaslaştırıcı bir uyaranın kolaylaştırıcı etkisi, serotonin benzeri bir maddenin etkisiyle gerçekleştirilebilir. Bu tür maddelerin etkisi, içlerinde bir aksiyon potansiyelinin gelişi sırasında gözlenen, kalsiyum iyonlarının sinir uçlarına normal girişini artırarak gerçekleştirilir (Schwarz ve diğerleri, 1983).
Sinaptik depresyon ekonomik bir süreçtir çünkü iletimi engellemek için hiçbir karmaşık bağlantı gerekmez . Tüm sinapslar bu özelliğe sahip olmasa da, yalnızca uyaran tarafından uyarılan sinapslar tarafından deneyimlenir . Bozulmamış bir organizmada, fizyolojik uyarılma seviyelerindeki bazı sinapslar oldukça güvenilirdir ve presinaptik liflerin her boşalmasıyla impulsları iletmeye devam ederler. Bu tür sinapslar genellikle bulunur.
birincil duyusal yollarda olduğu kadar yakın motor çıktıda, örneğin omuriliğin monosinaptik refleks yaylarında (Sherrington, 1906; Prosser ve Hunter, 1936; Spencer ve diğerleri, 1966c) ve nöromüsküler kavşaklarda. Bazıları yukarıda açıklanan diğer sinapsların güvenilirlik düzeyi düşüktür, iletme yeteneğini hızla kaybeder ve yavaş yavaş iyileşir. Bu nedenle, sinaptik iletim, geniş bir güvenilirlik ve kurtarma oranları yelpazesine sahiptir . Aslında, durum böyle olmasaydı , özellikleri farklı sistemlerde ve farklı hayvan türlerinde büyük farklılıklar gösteren davranış alışkanlığını açıklamak için sinaptik aktarım depresyonuna nasıl başvurulabileceğini hayal etmek zor olurdu (Hinde, 1970). Bunu akılda tutarak bile, belirli bir hayvanda bile tüm davranışsal alışkanlık durumlarında, tepkilerdeki azalmayı uygulamak için aynı hücresel mekanizmaların kullanıldığını varsayarak, bağımlılığın deneysel analizinin tamamlanmaya yakın olduğunu varsaymak için henüz çok erken. ve sensitizasyon (Horn, 1967; Sokolov ve diğerleri, 1967; Wickelgren, 1967b; Wall, 1970; Weight ve Erulkar, 1976). Tek hücreli organizmalarda, alışmaya benzeyen davranış değişiklikleri, neredeyse sinaptik mekanizmalara dayanmaz (Applewhite ve Gardner, 1971).
Alışkanlığa aracılık eden varsayılan sinir ağları
Yukarıdaki tartışmalarda, bir tür davranış değişikliğinin, yani alışkanlığın nöronal temelleri , elektroensefalogramdan elektriksel temaslara ve son olarak da moleküler düzeye kadar giderek daha temel düzeylerde incelenmiştir . Geriye gitmek daha zordur, çünkü daha düşük bir seviyedeki analizden çıkarılamayan yeni özellikler daha yüksek bir seviyede görünebilir. Ancak, bunu yaparken sadece mevcut bilgide boşluklar bulsak bile, farklı analiz seviyelerinde elde edilen sonuçları sentezlemeye çalışmakta fayda var.
Aşağıdaki tartışmada, bazı davranışsal veya fizyolojik tepkileri doğrudan kontrol eden nöronlardan birinin bir mikroelektrot tarafından kaydedildiği varsayılmaktadır . Böyle bir nöronun tekrarlanan bir uyarana yanıt vermeyi bıraktığını, ancak başka bir uyarana güçlü bir şekilde yanıt verdiğini varsayalım ve orijinal uyaranın eylemi bir süre durursa, o zaman tekrar sunulduğunda hücre ona yeniden yanıt vermeye başlar. Bu tür dinamikler, sinyali tekrarlanan bir uyaranla ileten yollarda otomatik olarak oluşturulmuş bir depresyon meydana gelirse gelişebilir (Şekil 1.11, A).
3-140

Resim: 1.11. Davranışsal alışmanın sinir ağlarının modelleri. A. Kayıtlı nöron R'deki yanıtın uyarana özgü zayıflaması için basit bir ağ. Stimulus S1, P -R yolu boyunca hücre üzerinde etki gösteren bir nöron kompleksini uyarır . S1 uyarısı tekrarlandığında, P hücresinin sinapsları yavaş yavaş uyarılmayı durdurur ve R hücresi bu uyarana yanıt vermeyi bırakır. Bununla birlikte, R hücresi , onu QR yolu boyunca uyaran S2 uyaranına yanıt verir . Bu yol anatomik olarak P-R'den bağımsızdır. B. Uyaran genellemesini gösteren bir ağ. Sinapsların sembolleri, Şekil 1'deki ile aynıdır. 1.10. Daha ayrıntılı açıklamalar için metne bakın. (Boynuz, 1967.)
Yukarıdaki şemada, S1 uyaranı, örneğin cilde hafif bir dokunuş, kayıtlı R nöronunda bir yanıta neden olur, P-R yolu üzerinden ona etki etmek. Dokunma tekrarlanırsa , P hücresinin terminallerindeki sinaptik depresyon nedeniyle R'ye iletim engellenir . güçlü bir hücresel tepki eşlik eder. Sinaptik depresyon devam ettiği sürece , S1 uyarısı R hücresinden bir yanıt elde edemeyecek , ve sonuç olarak, karşılık gelen davranışsal tepki, yani sinaptik depresyonun süresi, bu uyarana yanıt verilmemesi gereken hafızanın tutulma süresini belirler. Bu nedenle, ilk uyaranın belirli bir süre sonra bir tepki uyandırma yeteneği, sinaptik depresyonun kaybolma hızına bağlı olacaktır . Bu işlem tamamlandığında ve reaksiyon geri geldiğinde hafıza kaybolur. Güçlü bir ekstra uyaranın , bastırılmış bir sinaps aracılığıyla iletimi yeniden sağlayarak "unutmayı" nasıl tetiklediği yukarıda tartışılmıştır (bkz. Şekil 1.10).
Bu nedenle, tepkinin kademeli olarak baskılanması ve zaman içinde iyileşmesi , birinci ve ikinci uyaranların uyandırdığı impulsların kayıtlı hücreye anatomik olarak bağımsız yollardan ulaştığı varsayılarak açıklanabilir . Bu varsayım sebepsiz yere yapılmaz. Uyaranın belirli özelliklerinin, belirli nöron gruplarının aktivitesini uyardığına dair çeşitli hayvanlar üzerinde elde edilen oldukça ikna edici veriler vardır . Örneğin, retina boyunca bir yönde hareket eden bir çizgi serebral korteksteki bir grup nöronu uyarırken, ters yönde hareket eden bir çizgi diğerini uyarır (Hubel, 1982). İşitme sisteminde birçok nöron belirli bir frekanstaki sese seçici olarak yanıt verir (Whitfield, 1967); bu nedenle, bir sesin frekans bileşenlerindeki bir değişiklik, uyaran tarafından ateşlenen nöron setinde bir değişikliğe neden olacaktır. İşitme sistemindeki bazı nöronlar , yalnızca belirli bir frekans veya yoğunluktaki seslerde ateşlenir (Goldberg ve Greenwood, 1966). Bu durumda, tek başına ses yoğunluğundaki bir değişiklik, başka bir nöron popülasyonunu uyarmak için yeterli olacaktır .
Alışkanlığın nöral mekanizması, bir örneği Şekil 1'de gösterilen uyaranların genelleştirilmesini de açıklamalıdır. 1.3. Başlangıçta, hücre 1000 ve 950 Hz'lik tonlara yanıt verdi. 1000 Hz'lik bir tona yanıt söndükten sonra, 950 Hz'de yanıt da kayboldu. Bu etki, Şekil 1'de gösterildiği gibi yol örtüşmesi ile açıklanabilir. 1.11, B. S1 uyaranının (ton 1000 Hz) nöron 1-4'ü uyardığı ve bunun da R hücresini uyardığı varsayılmaktadır. S2 uyaranının (950 Hz) ayrıca hücre 7'yi de uyardığı, ancak nöronları içeren bir yol aracılığıyla 3 -6. Her iki uyaran da 3. ve 4. hücreleri aktive eder. Ayrıca, 7. ve 8. hücrelerin ateşlenmesini sağlamak için üç veya daha fazla girdi nöronunun aktivasyonunun gerekli olduğu varsayılır. 1000 Hz'lik bir tonun tekrar tekrar sunumu üzerine, girişlerindeki sinaptik depresyon nedeniyle 1-4 hücrelerinin yanıtları azalır. Hücre yanıtları 7 ve R da zayıflıyorlar. Şimdi 950 Hz'lik bir ton sunun. 5 ve 6 numaralı hücreler ateşlenir, ancak 3 ve 4 numaralı hücreler giriş sinapsları baskılandığı için ateşlenmez. Sonuç olarak, hücre 8'in uyarılması için gerekli koşullar yoktur, çünkü minimum aktif giriş sayısı ile aktif hale getirilmemiştir. Hücre R, bu nedenle bir deşarj oluşturmaz.
Tek başına sensorimotor yolaklarda otomatik olarak oluşturulan iletim depresyonu ile açıklanamayan veriler vardır. İnsanlar üzerinde araştırma yapan Sokolov (1960), düzenli olarak tekrarlanan bir uyarana yöneltici tepkinin sönmesinden sonra , dizideki uyaranlardan Ding atlanırsa tepkinin yeniden ortaya çıktığını bulmuştur. Sokolov (Sokolov, 1965) 3'ü tanımladı

Resim: 1.12. Bir nöronun kaçırılan bir uyarana yanıt verebildiği bir sistem. G hücresinin aktivitesinin, P ve CB hücrelerinin aktiviteleri arasındaki farkla orantılı olduğu bir Fark saptama devresi . metinde bu tür reaksiyon türlerinden biri tartışılmaktadır. Sinapsların tanımları, Şekil 1'deki gibi. 1.10. (Boynuz, 1967.)
ayrıca tavşanın hipokampüsünde uyarı kaçırıldığında yanıt veren tek hücreler.
bu zamana bağlı değişiklikleri ve aynı zamanda davranışsal düzeyde yönlendirme tepkisinin benzer tezahürlerini açıklamak için, otojenere depresyona ek olarak bazı ek süreçlerin getirilmesi gerekli görünmektedir. Böyle bir süreç , farklılıkları tespit eden bir sistemin çalışmasını gerektirmelidir . Bu sistemin çalışma prensibi Şekil 1 de gösterilmiştir. 1.12A.P ve J hücrelerinin bir duyusal uyaran tarafından aynı anda aktive edildiğini ve her birinin belirli bir zaman aralığında n sivri uç ürettiğini varsayalım.Kilitleyici girdiler hassas bir şekilde dengelenmiştir ve J hücresi sessizdir.Hücre I de aynı şekilde sessizdir. nedenler ve G hücresinden beri J veya H hücreleri tarafından uyarılmaz , aynı zamanda sessizdir.
Farklılıkları saptamak için kullanılan devre , Şekil 1'de gösterilen devreye dahil edilmiştir. 1.12, B. Tavşanların ve kedilerin hipokampüsünde ve görsel korteksinde kayıtlı nöronların özelliklerinin atfedildiği K hücresini içerir (Horn, 1962; Kopytova ve Rabinovich, 1967; Vinogradova, 1970; Brown, 1982). Bu hücrelerin deşarjları aşağıdaki uyaranlarla düzenli olarak tetiklenir . Bu fenomenin mekanizması bilinmemektedir, ancak sonucu, hücrelerin birkaç ekstrapolasyon deşarjı, yani beklenen görünüm anında uyarıyı sunmadan hücrede meydana gelen deşarjlar verme yeteneğidir .
Yeni bir uyaranın sunulması üzerine, P hücresi boşalır ve R hücresi, PFR yolu boyunca aktive edilir (Şekil 1.12, B). G hücresi sessizdir çünkü K hücresindeki deşarj P hücresinin deşarjına eşittir, tekrar uyaran uygulandığında R hücresinin tepkisi azalır . Bir dizide bir uyaran kaçırıldığında, K hücresi bir ekstrapolasyon deşarjı verir. Hücre J ayrıca boşalır, çünkü K hücresi onu uyarır ve sessiz P hücresinin inhibitör girdisi yoktur. G hücresi , F ve R hücreleri gibi boşalır ve böylece kaçırılan uyarana yanıt verir.
Yorumlar
Yukarıda tartışılan nöronal mekanizmalar , bir uyaranın tekrar tekrar sunulmasından kaynaklanan davranış değişikliklerinin birçok yönünün altında yatıyor olabilir. Belirli bir uyarana yanıt vermenin gerekli olmadığına dair belleğin sinirsel temeli , başlangıçta tekrarlanan uyaran tarafından uyarılan yoldaki otojenleştirilmiş bir iletim depresyonu olabilir; bu etkisiz hale getirilmiş yol, o uyaranın nöral modelini temsil eder. Ancak bu tür sistem ve mekanizmalar, zaman refleksini açıklamak için başvurulması gereken sistem ve mekanizmalarla birlikte, davranış değişikliğinin tüm yönlerini yeterince sağlayabilir mi? Muhtemelen değil. Hinde'nin (1954a, b) ispinozların (Fringilla coelebs) saldırı tepkilerine ilişkin çalışmasının sonuçları dikkate alındığında zorluklar belirginleşir. bazı avcılarda. Bir yırtıcı hayvanın - doldurulmuş bir baykuş - sürekli varlığıyla, reaksiyon tamamen ortadan kalktı. Baykuş 24 saat sonra tekrar gösterildiğinde tepki ilk seferden daha zayıftı. Davranıştaki bu değişiklik, duyusal girdiyi düşmeye tepkiyi kontrol eden nöral sistemlere bağlayan yollardaki kendiliğinden oluşan iletim depresyonu ile açıklanabilirse, sonraki verileri hesaba katmak için ek bir sürecin varsayılması gerekir (Hinde, 1960). Bu deneyler iki uyaran kullandı, bunlardan biri baykuş, diğeri köpekten daha etkiliydi. Uyarıcılar, aralarında 24 saat arayla iki farklı deneyde sunuldu. Uyaranların sunum sırası ve ilk sunumun süresi değişmiştir. Çoğu durumda, uyaranın ikinci sunumundaki reaksiyonun gücü ile değerlendirilen iyileşme, ilk sunum ne kadar uzunsa, o kadar zayıftı . Bununla birlikte, güçlü bir uyaranın ardından zayıf bir uyaranın geldiği durumlarda , ters etki gözlendi: baykuşun ilk sunumu ne kadar uzunsa, köpeğe sonraki tepki o kadar güçlüydü. Böylece, bir durumda, birinci sunulan uyaranın ikincisi üzerinde uzun süreli azaltıcı bir etkisi gözlenirken, diğer durumda artırıcı bir etki gözlemlendi. Bu iki etki tek bir eksiltme işlemiyle açıklanamaz. Bu nedenle, Hinde (1960, 1970), bu durumda iki sürecin söz konusu olduğunu öne sürdü - azalma ve artım. Bu bölümde açıklanan fizyolojik araştırmalar, her iki süreç de hayvan tekrarlanan uyaranlara maruz kaldığında ortaya çıksa da artımlı süreçten ziyade azalma süreci, alışma ve bunun düzenlenmesi için koşullarla ilgilendi.
Alışmanın bazı yönlerinin nöral analizinde son yirmi yılda önemli ilerlemeler kaydedilmiştir ve coşku dalgası, araştırmacıları alışma ve duyarlılaştırma mekanizmaları açısından çeşitli ilişkisel öğrenme türlerini açıklamaya teşvik edecek kadar güçlü olmuştur (Horn, 1971a; Hawkins ve Kandel, 1984) . Bu başarılar bir dizi faktöre bağlıydı. Bunlardan biri, bu süreci analiz etmek için teknik yöntemlerin geliştirilmesi ve kullanılabilirliğidir . Diğer bir faktör, incelenen sorunun doğasında yatmaktadır . Alışma, çeşitli koşullar altında doğru bir şekilde tanımlanabilen ve gözlemlenebilen bir davranış değişikliğidir . Ek olarak, uyaran ve tepki genellikle zamanla yakından ilişkilidir. Birçok hayvanın sinir sisteminde ve hatta sinir sisteminin izole edilmiş kısımlarında davranış değişikliklerine tam parametrik benzerlik gösteren nöronal kaymalar gözlemlenmiştir. Tüm bu faktörlerin kombinasyonu, bu tür davranışları nörobiyolojik laboratuvarlarda incelemeyi mümkün kıldı. Bununla birlikte, en düşük nöron aktivitesi seviyesinde analiz için erişilebilir davranış biçiminin seçimi, çalışmanın başarısı için en temel unsurdu.
Yaklaşık 15 yıl önce, diğer öğrenme biçimlerini açıklayacak bilgi depolamanın nöral temelini keşfetme olasılığı yeterince cesaret verici değildi. Bunun nedenleri açıktır. Çoğu durumda, uyaran başlangıçta belirli bir tepkiye neden olmaz, ancak bu özelliği öğrenme sürecinde kazanır. Bu durumlarda, uyaran ve tepki zaman içinde yakından ilişkili olmayabilir: tepkiler öğrenme sırasında değil, daha sonra ortaya çıkabilir; yeni bir davranışın oluşumu, olumlu veya olumsuz pekiştirmenin kullanılmasını gerektirir ve bu davranış karmaşık olabilir ve hayvanın entegre bir hareketini gerektirebilir; son olarak, tepkinin doğasını belirlemede geçmiş deneyimler büyük önem taşıyabilir. Bu nedenle, bilgi depolamanın temelini oluşturan beyinde değişiklikler oluşturmaya yönelik girişimlerin büyük zorluklarla karşılaşması şaşırtıcı değildir . Öğrenme ve hafızanın temelleri üzerine yapılan çalışmaların davranış bilimleri ve beyin bilimleri arasındaki arayüzde yatan problemler ortaya koyduğu düşünüldüğünde, bu güçlükler daha da artar. Davranış araştırmacıları bunu analiz etme yeteneğine sahiptir, ancak sinirsel mekanizmaları nadiren inceleyebilirler ve mekanizmaları inceleyebilen sinirbilimciler, davranış değişikliğinin meydana geldiği koşulları nadiren belirleyebilir ve tanımlanmış mekanizmaların davranış değişikliğini tam olarak belirleyebilirler. ilişkilidir. Bu nedenlerden dolayı, 1960'larda öğrenme ve hafıza üzerine yapılan biyokimyasal çalışmalarda büyük ölçüde başarı umutları büyük ölçüde gerçekleşmedi (Horn, 1971b).
, bir ışık parlaması gibi başlangıçta kayıtsız bir uyaranın ödül veya ceza konusunda uyarmaya başladığı klasik koşullu reflekslerin oluşumu için gerekli olan hafızanın analizinde önemli ilerleme kaydedildi . Bu çalışmalar birkaç mükemmel incelemede gözden geçirildi (Voronin, 1976; Woody, 1982; Thompson ve diğerleri, 1983; Alkon, 1984), bu yüzden burada onlar üzerinde durmayacağız. Bununla birlikte, görünüşe göre deneysel analize konu olabilecek başka bir öğrenme türü daha var - evcil tavukta (Gallus domesticus) baskılama. Böyle bir iddia için yeterli gerekçeler var.
Baskının özellikleri
doğumdan birkaç saat sonra yürüyebilen ve koşabilen birçok hayvan türünün yavrularında doğaldır . Bu "olgun doğmuş" türler, doğumda neredeyse çaresiz olan ve günlerce, haftalarca veya aylarca bu şekilde kalan "olgunlaşmamış" türlerden farklıdır . Koyun, keçi, kobay ve bazı antiloplar gibi bazı memelilerde damgalama meydana gelir , ancak bu davranış özellikle olgun (kuluçka) kuşlarda kapsamlı bir şekilde incelenmiştir (Spalding, 1873; Heinroth, 1910; Lorenz, 1935, 1937; Hinde, 1962; Bateson, 1966; Sluckin, 1972; Hess, 1973).
Kuluçkadan çıktıktan kısa bir süre sonra damızlık kuşların civcivleri annelerini takip etmeye başlar. Ancak aşağıdaki tepki sadece gerçek anneden kaynaklanmaz. Örneğin, evcil tavuklar UR'ye çok çeşitli nesnelere, özellikle hareketli olanlara yaklaşır. Civcivler belirli bir nesneye uzun süre maruz kalırlarsa, gerçek bir anneye olan bağlılığa benzer bir bağlanma geliştirirler . Civcivler tanıdık bir nesnenin yakınındayken zevk belirtileri gösterirler: sessizce ciyaklarlar, küçük yiyecek parçalarını gagalarlar ve nesne hareket ederse civcivler onu takip eder. Dahası, diğer görünür nesnelere, damgalama gerçekleşmeden önce yapacakları gibi yaklaşmak yerine, korku ve hatta panik belirtileri göstererek onlardan aktif olarak kaçınmaya başlarlar . Bu davranış , civcivlerin "annenin" işaretlerini hatırladığını gösterir.
Civcivlerin çeşitli nesnelere yaklaştığı dönem genellikle "hassas dönem" olarak adlandırılır. Bu dönemden önce tavuklar çok uyurlar ve uyanık olduklarında hareketleri zayıf bir şekilde koordine edilir. Bu nedenle, hassas dönemin başlangıcı gelişme derecesi ile sınırlı olabilir - lokomotor aktivitenin olgunlaşmamışlığı ve yumurtadan çıktıktan sonraki ilk saatlerde uyuşukluk. Yeni nesnelerden kaçınma, hassas dönemin sona erdiğini gösterir.
Yavru damgalama, genç kuşların bir anneye veya yapay bir ikameye bağlanma geliştirdiği süreçtir. Cinsel eş seçimini etkileyen cinsel damgalama daha geç ortaya çıkar (Schutz, 1965; Vidaî, 1980). Cinsel damgalama, genç bir kuşun başkalarıyla, özellikle yavru arkadaşları (kardeşleri) ve annesiyle iletişim deneyiminden etkilenir . Cinsel damgalamanın sonuçları genellikle ancak daha sonra, haftalar veya aylar sonra, kuş cinsel olgunluğa eriştiğinde keşfedilir . Bundan önceki dönemde kuş, sinir sisteminde iz bırakabilen çeşitli etkilere maruz kalır. Buna karşılık, civciv damgalamada civciv , nesne ona çarptığı anda doğrudan yaklaşarak veya onu takip ederek tepki verir. Etkinin yakınlığı , civciv damgasını öğrenme sürecinin nöral mekanizmalarını analiz etmek için çekici bir nesne haline getirir.
Deney hayvanı ve eğitim koşulları seçilirken bir dizi faktör dikkate alınmalıdır . Olgun memelilerde nesne bağlanmasının gelişimi ile ilgili veriler nispeten azdır (Sluckin, 1972). Memelilerde damgalamanın belirgin tezahürleriyle, tercih gelişimi büyük ölçüde koku alma uyaranlarına bağlı görünmektedir (Carter ve Magg, 1970; Porter ve Etscorn, 1974) ve koku alma uyaranlarının kontrol edilmesi zordur. Örneğin işitsel uyaranlar açılıp kapatılabiliyorsa koku alma uyaranları uzun süre kalır. Ayrıca genç memeliler anneleri tarafından beslenirler, bu nedenle onun kokusuna karşı bir bağlılık geliştirebilirler. Bu faktörler , olgun doğmuş memelilerde imprinting deneysel çalışma olasılığını ciddi şekilde sınırlar . Bu nedenle , bu hayvanlar, nöronal damgalama mekanizmalarının analizi için ilk etapta seçilmemelidir.
Kuluçka kuşlarının pek çok civciv işitsel veya görsel uyaranlara kolayca baskı oluşturabilir. Baskılama çalışmalarında işitsel uyaranların bazı eksiklikleri vardır. Örneğin, yaban ördeği civcivleri yumurtadan çıkmadan önce ses çıkarmaya başlarlar (Gottlieb, 1971), dolayısıyla bu kuşların bireysel deneyimlerini işitsel alemde kontrol etmek zordur. Sonunda araştırma için evcil civcivleri seçtik çünkü davranışları iyi anlaşıldı, beyinleri deneysel manipülasyon için yeterince büyük, yumurta kesesi yumurtadan çıktıktan sonra saatlerce besin sağlıyor ve onları elde etmek zor değil. Tavuklarda damgalamayı incelemenin temel avantajı , belirli koşullar altında gördükleri ilk görsel nesnenin özelliklerini sabitlemeleridir . Deneyci, böyle bir nesne ortaya çıkana kadar tavukları karanlıkta yetiştirerek, görme yoluyla alınan bilgilerin öğrenmeden önce beyinde bulunmadığından emin olabilir . Elbette bir tavuğun beyni, deneyimin iz bıraktığı bir tabula rasa değildir, ancak diğer hayvanların beyniyle kıyaslandığında buna en iyi yaklaşan beyindir .
Özet
Beyindeki bilgi depolamanın nöral temellerine olan ilgi uzun bir geçmişe sahiptir. Son yirmi yılda, alışkanlığın bazı yönleriyle ilişkili izlerin depolanmasının analizinde kayda değer ilerlemeler oldu . Alışkanlık , tekrarlanan bir uyarana verilen davranışsal veya fizyolojik tepkinin, uyaran bir pekiştireç ile ilişkili değilse kademeli olarak ortadan kalktığı bir öğrenme şeklidir. Bu bellek olumsuzdur; varlığı, bir uyarana tepki olmamasından anlaşılır. Alışkanlık , tüm evrimsel seviyelerdeki hayvanlarda bulunmuştur. Sinir sisteminin izole edilmiş bölümleri tarafından kontrol edilen reaksiyonlar için, parametreleri sağlam bir sinir sistemi olan hayvanlardan daha yüksek olabilmesine rağmen, alışkanlık da karakteristiktir. Bu veriler, davranışsal alışkanlığın, duyu reseptörleri ve yanıt kontrol eden nöronlar arasındaki yollar boyunca sinapslarda otojenleştirilmiş iletim depresyonuna bağlı olduğunu göstermektedir . Belirli bir uyaran tarafından etkinleştirilen belirli bir nöron grubundaki iletim depresyonu, bir yanıtın yokluğunda ifade edilen hafıza izinin temeli gibi görünüyor.
Diğer öğrenme biçimlerinde, yalnızca mevcut bir yanıtın kaybı değil, yeni bir yanıtın geliştirilmesi veya mevcut bir yanıtın değiştirilmesi söz konusudur . Bu öğrenme biçimlerinde bilgi depolamanın nöronal temeli üzerine yapılan çalışmada, "azalan" davranış veya alışkanlık çalışmasında olduğundan daha karmaşık problemler ortaya çıkar. Bunun sebepleri değerlendirilir. Ayrıca , damgalama çalışmasının, hafızanın nöronal temellerini analiz etme olasılığı için neden umut verdiği de gösterilmiştir. Evcil tavukta civciv baskısının kısa bir açıklaması verilmiştir.
EDEBİYAT
D. L, Kalsiyumla inaktive edilmiş potasyum çevreleri: bir biyofiziksel hafıza izi, Science, 1984, 226, 1037-1045.
Applewhite PB, Gardner FT Theory of protozoan habituation, Nature new Biol., 1971, 230, 285-287.
Bateson PPG Baskının özellikleri ve bağlamı, Biol. Rev., 1966, 41, 177-220.
Bell C., Sierra G., Buendia N., Segundo JP Mezensefalik retiküler formasyondaki nöronların duyusal özellikleri, J. Neurophysiol., 1964, 27, 961-987.
Berger H. Uber das Electrenkephalogramm des Menschen. Zweite Mitteilung, J. Psychol. Neurol., 1930, 40, 160-179.
Brindley GS Değiştirilebilir sinapsların sınıflandırılması ve koşullandırma modellerinde kullanımları, Proc. R. Soc., 1967, B168, 361-376.
Kedi hipokampal nöronlarının aktivitesinde geçici bir hafıza için Brown MW Kanıtı, J. Physiol., Lond., 1982, 328, 42-3P.
Bruner J., Tauc L. Aplysia'da sinaptik düzeyde alışkanlık, Nature, Londra., 1966, 210, 37-39.
Bryant SH Kalamar yıldız gangliyonunun proksimal sinapsının işlevi, J. gen. Physiol., 1959, 42, 609-616.
Buchwald JS, Halas ES, Schramm S. Tekrarlanan kutanöz stimülasyona götüren ünite yanıtlarında progresif değişiklikler, J. Neurophysiol., 1965, 28, 200-216.
Bullock TH Özellikleri - kalamarın yıldız ganglionundaki tek bir sinapsın, J. Neurophysiol., 1948, 11, 343-364.
Bullock TH, Hagiwara S. Kalamarın dev sinapsından hücre içi kayıt, J. gen. Physiol., 1957, 40, 565-577.
Cajal S. R. In Histologie du systeme neurux de l'homme et des vertebres, 1911, Cilt. 2, s. 886-890. Malone, Paris. (Yeniden basım 1955, Histologie du systeme neerux. Instituto Ramon y Cajal, Madrid.)
Carter CS, Marr JN Gine domuzu Anim'de koku alma damgası ve yaş değişkenleri. Behav., 1970, 18, 238-244.
Castellucci VF, Kupfermann L, Pinsker H., Aplysia, Science, 1970, 167, 1445-1448'de Kandel ER Solungaç geri çekme refleksinin alışma ve dishabitüasyon nöronal mekanizmaları .
Castellucci VF, Carew TJ, Kandel ER Aplysia californica'nın solungaç çekme refleksinin uzun vadeli alışkanlığının hücresel analizi , Science, 1978, 202, 1306-1308.
Cynader M., Berman N. Monkey superior colliculus'un alıcı alan organizasyonu , J. Neurophysiol., 1972, 35, 187-201.
Descartes R. İçinde: Descartes'ın temel eserleri (transi. L. Blair). Bantham Books, New York, 1961.
Dräger UC, Hubel DH Görsel uyarıya yanıt ve fare üst kollikulusunda görsel, işitsel ve somatosensoriyel girdiler arasındaki ilişki, J. Neurophysiol., 1975, 38, 690-713.
Dudel J. 5-hidroksitriptaminin kerevit nöromüsküler kavşağı üzerindeki kolaylaştırıcı etkileri, Naunyn-Schmiedebergs Arch. tecrübe. Yol. Pharmak., 1965, 249, 515-528.
Ectors L. Etüde de l'activite electrique du cortex serebral chez le Lapin non narcotise ni curarise, Arch. int. Physiol., 1936, 43, 267-298.
Foster M. Bir fizyoloji ders kitabı. Macmillan, Londra, 1897.
Freud S. Bilimsel bir psikoloji projesi 1895. İçinde: Sigmund Freud'un tüm psikolojik çalışmaları (ed. J. Strachey), 1966, Cilt. 1, Hogarth Press, Londra .
Glaser EM, Whittow GC Lokalize soğumaya adaptasyon için sıcak bir ortamın tutulması, J. Physiol., Lond., 1957, 136, 98-111.
Goldberg JM, Greenwood DD Kedinin dorsal ve posteroventral koklear çekirdeklerinin nöronlarının uzun süreli akustik uyaranlara tepkisi, J. Neurophysiol., 1966, 29, 72-93.
Gordon B. Cat superior colliculus'un derin katmanlarındaki alıcı alanlar, J. Neurophysiol ., 1973, 36, 157-778.
Gottlieb G. Kuşlarda tür tanımlamasının gelişimi: algının doğum öncesi belirleyicilerine yönelik bir araştırma. Chicago Press Üniversitesi, Chicago, 1971.
Griffin JP, Pearson JA Farede fleksör refleks alışkanlığı, J. Physiol., 1967, 190, 3-5P.
Griffith JS Belleğin doğasına ilişkin bir teori, Nature, Londra., 1966, 211, 1160-1163.
Grüsser-Cornehls U., Grüsser OJ, Bullock TH Unit, kurbağanın tektumunda hareketli ve hareketsiz görsel uyaranlara tepkiler, Science, 1963, 141, 820-822.
Hagiwara S., Tasaki /. Mürekkep balığının dev sinapsı boyunca dürtü aktarım mekanizması üzerine bir çalışma, J. Physiol., Lond., 1958, 143, 114-837.
Hawkins RD, Kandel ER Basit öğrenme biçimleri için bir hücre-biyolojik alfabe var mı? Psikolog. Rev., 1984, 91, 375-391.
Hebb D. 0. Davranışın organizasyonu, John Wiley, New York, 1949.
Heinroth O. Beitrage zur Biologie, namentlich Ethologie ve Psychologie der Anatiden, Verh. 5. int. Om. Kongr., 1910, 5, 589-702.
Held H. Beitrage zur Struktur der Nervenzellen und ihren Fortsâtze, Arch. Anat. fizik. Lpz., 1897, 204, Ek. 273.
Hess EH Damgalama: erken deneyim ve bağlanmanın gelişimsel psikobiyolojisi. Van Nostrand Rheinhold, New York, 1973.
(Fringilla coelebs) yıldırma davranışında gösterildiği gibi, kısmen doğuştan gelen bir tepkinin gücündeki değişiklikleri yöneten Hinde RA Faktörleri. I. Yanıtın doğası ve seyrinin incelenmesi, Proc. R. Soc, 1954a, B142, 306-331.
(Fringilla coelebs) yıldırma davranışında gösterildiği gibi, kısmen doğuştan gelen bir tepkinin gücündeki değişiklikleri yöneten Hinde RA Faktörleri. II. Yanıtın azalması, Proc. R. Soc., 1954b, B142, 331-358.
İspinozun (Fringilla coelebs) mobbing davranışında gösterildiği gibi, kısmen doğuştan gelen bir tepkinin gücündeki değişiklikleri yöneten Hinde RA Faktörleri. III. Kısa vadeli ve uzun vadeli artan ve azalan etkilerin etkileşimi, Proc. R. Soc., 1960, B 153, 398-420.
Hinde RA Damgalama sorununun bazı yönleri, Symp. hayvanat bahçesi Soc., Londra., 1962, 8, 129-138.
Hinde RA Davranışsal alışkanlık. İçinde: Nöral aktivite ve davranışta kısa vadeli değişiklikler (eds. G. Horn ve R.A. Hinde)', 1970, s. 3-40, Cambridge University Press, Cambridge.
Hobson JA, Scheibel A. B. (editörler) Beyin sapı çekirdeği: sensorimotor entegrasyon ve davranışsal durum kontrolü, Neurosci. Res. programı. Bull., 1980, 18, 1-173.
Holmgren B., Frenk S. Nöronal seviyede inhibitör fenomenler ve “alışkanlık”, Nature, Londra., 1961, 192, 1294-1295.
Horridge GA, Scholes JH, Shaw S. t Tunstall J. Çekirgenin optik lobundaki ve beynindeki tek nöronlardan hücre dışı kayıtlar. İçinde: Böcek Merkezi sinir sisteminin fizyolojisi (eds. JE Treherne ve JWL Beament). Academic Press, Londra, 1965.
Horn G. Algının bazı nöral korelasyonları. İçinde: Biyolojide bakış açıları (eds. JD Carthy ve CL Duddington), 1962, Cilt. 1, s. 242-285. Butterworth, Londra.
Horn G. Seçici algının fizyolojik ve psikolojik yönleri. İçinde: Davranış çalışmasında gelişmeler (eds. D. Lehrman, R. A. Hinde ve E. Shaw), 1965, Cilt. 1, s. 155-215, Academic Press, New York.
Horn G. Nöronal alışma mekanizmaları, Nature, Londra., 1967, 215, 707-711.
Boynuz G. Yenilik, dikkat ve alışkanlık. İçinde: Nörofizyolojide bir kavram olarak dikkat (editörler CR Evans ve T. Mulholland), 1969, s. 230-246. Tereyağı değerinde, Londra.
Horn G. Yeni ve tekrarlanan uyaranlara davranışsal ve hücresel tepkiler. İçinde: Davranışın nöral kontrolü (eds. RE Whalen, RF Thompson, M. Verzeano ve NM Weinberger), 1970a, s. 103-127, Academic Press, New York.
Horn G. Nöronal aktivite ve davranıştaki değişiklikler. İçinde: Nöral aktivite ve davranışta kısa vadeli değişiklikler (eds. G. Horn ve R.A. Hinde), 1970b, s. 567-606.Cambridge University Press, Cambridge.
Boynuz G. Alışkanlık ve hafıza. In: Biology of memory (ed. G. Âdâm), 1971a, s. 267-284, Akademiai kiado, Budapesi.
Boynuz G. Merkezi sinir sisteminde deneyimle ilişkili biyokimyasal morfolojik ve işlevsel değişiklikler , Activitas nervosa superior, 1971b, 13, 119-130.
Horn G., Hili RM Tavşanların beyin sapındaki nöronların duyusal uyaranlara tepkisinin alışması, Nature, Londra., 1964, 202, 296-298.
Horn G., Hili RM Tavşanın superior colliculus ve altındaki tektotegmental bölgelerindeki birimlerin duyusal stimülasyonuna duyarlılık Exp. Neurol., 1966, 14, 199-223.
Horn G., Rowell CHF Çekirgelerin tritocerebrnamındaki görsel nöronların davranışındaki orta ve uzun vadeli değişiklikler, J. exp. Biol., 1968, 49, 143-170.
Horn G., Wright MJ Kalamar yıldız ganglionundaki iletim hatasının özellikleri: basit bir alışma sistemi üzerine bir çalışma, J. exp. Biol., 1970, 52, 217-231.
Hubel DH İlk görsel korteksin keşfi . 1955-1978, Nature, Londra., 1982, 299, 515-524.
Hughes GM, Tauc L. Aplysla depilans'taki iki dev sinir hücresinin anatomik ilişkilerinin eledrofizyolojik bir çalışması, J. exp. Biol., 1963, 40, 469-486.
Humphrey G. Le Chatalier'in kuralı ve Helix albolabris, Psychol'da alışkanlık ve dehabitüasyon sorunu . Forsch., 1930, 13, 113-127.
Humphrey G. Canlı sistemle ilişkisinde öğrenmenin doğası. Harcourt, New York, 1933.
Jassik-Gerschenfeld D. Kedinin superior kollikulusunda uyandırılan somatik kaynaklı aktivite, Exp. Neurol., 196s, 10, 104-118.
Katz B. Nöral verici maddelerin salınması. Liverpool University Press, Liverpool, 1969.
Katz B., Miledi R. Nöromüsküler iletim sırasında kalsiyum etkisinin zamanlaması , J. Physiol., Londra., 1967, 189, VEVMI44.
Katz B., Miledi R. Presinaptik terminallerde tetrodotoksine dirençli aktivite, J. Physiol., Lond., 1969, 203, 459-487.
Kelly RB, Deutsch JW, Carlson SS, Wagner JA Biochemistry of neuro-transmiter release, A. Rev. Neurosci., 1979, 2, 399-446.
Klein M., Shapiro E., Kandel ER Ca 2+ akımının modülasyonunun sinaptik plastisitesi , J. exp. Biol., 1980, 89, 117-157.
Knott JR, Henry CE İnsan elektroensefalogramının alfa ritminin bloke edilmesinin şartlandırılması, J. exp. Psychol., 1941, 28, 134-144.
Kopytova F. V., Rabinovich M. G. Zamana şartlandırılmış refleksin mikroelektrot çalışması, Zh. daha yüksek, gergin deyat., 1967, 17, 1023-1033.
Krasne F.B., Roberts A. Pikrotoksin tarafından indüklenen inhibisyondan salınma sırasında kerevitin kaçış tepkisinin alışması, Nature, Lond., 1967, 215, 769-770.
Kupfermann L, Kandel ER Aplysia'nın abdominal ganglionunun aracılık ettiği davranışsal bir tepkinin Sinir Kontrolleri , Science, 1969, 164, 847-850.
Kupfermann L, Pinsker H., Castellucci V., Aplysia, Science, 1970, 167, 1743-1745'te Kandel ER Neuronal, solungaç geri çekme refleksinin alışma ve dishabituasyon bağıntıları.
Lashley KS Engram arayışında, Symp. sos. tecrübe. Biol., 1950, 4, 454-482.
terminaller, Proc. R. Soc., 1973, B183, 421-425.
Miledi R., Slater CR Kalsiyumun kalamardaki nöronal sinapslar üzerindeki etkisi, J. Physiol., Londra., 1966, 184, 473-498.
Milner PM Hücre meclisi: Mark II, Psychol. Rev., 1957, 64, 242-252.
Pavlov IP Serebral hemisferlerin çalışması üzerine dersler. Tamamlamak
anestezi uygulanmamış kedi, Am. J. Physiol., 1937, 119, 186-196.
Koberts MVB Solucan Lumbricus terrestris L. II'nin dev lif refleksi . Yorgunluk, J. exp. Biol., 1962, 39, 229-237.
Rowell CHF Böcek Merkezi sinir sisteminde artan ve azalan süreçler. İçinde: Nöral aktivite ve davranışta kısa vadeli değişiklikler (eds. G-Horn ve R.A. Hinde), 1970, s. 237-280, Cambridge University Press, ambridge.
Rowell CHF, Noth G. Bir böcek beynindeki tek sinir hücrelerinin tepkisinde davranış bozukluğu ve uyarılma, J. exp. Biol., 1968, 49, 1.71-183.
Sharpless S., Jasper H. Uyarılma tepkisinin alışması, Brain, 1956, 79 F 655-680.
Shepherd GM Beynin sinaptik organizasyonu (2. baskı). 1979, Oxford University Press, Oxford.
Bazı omurilik sinirlerinin arka köklerinin liflerinin periferik dağılımının incelenmesinde Sherrington CS Deneyleri, Phil. Trans. R. Soc., 1898, 190, 45-186.
Sherrington CS Sinir sisteminin bütünleştirici etkisi. 1906, Memur, Londra.
Schutz DP Duyusal kısıtlama, Academic Press, New York, 1965.
Schwarz JH, Bernier L., Castelluci VF, Palazzolo M., Saitoh T., Stapleton A. „ Kandel ER Aplysia'da kısa süreli hassaslaştırma için belleğin zaman sürecini hangi moleküler adımlar belirler ? Cold Spring Harb Symp. miktar Biol.,. 1983, 48, 811-819.
Sluckin W. Damgalama ve erken öğrenme. Methuen, Londra, 1972.
Sokolov TR Nöronal modeller ve yönlendirme refleksi. İçinde: Merkezi sinir sistemi ve davranış (ed. MAB Brazier), 1960, s. 187-276. Josiah Macy Jr Vakfı, New York.
Sokolov EH Algı ve koşullu refleks. Ed. Moskova Devlet Üniversitesi, 1958.
Sokolov E. N. Tek bir birim seviyesinde inhibe edici şartlandırılmış refleks, Proc. XIII int. Kongre Physiol., 1965, 4, 340-343.
Sokolov EH, Arakelov GG, Levinson LB Gastropod yumuşakça Limnea stagnatis'in kendiliğinden olmayan nöronlarının tekrarlayan elektriksel uyaranlara, Zhurn'a "Bağımlılığı". evrimsel biyokimya fiziol., 1967, 3, 147-153.
Spalding D. A. Instinct, genç hayvanlar üzerinde orijinal gözlemlerle, Macmillan's Mag., 1873, 27, 282-293. [1954'te Br'de yeniden basılmıştır. J. Animasyon Davranış, 2, 2-11.]
Spencer WA, Thompson RF, Neilson DR, Jr. Akut spinal kedide fleksiyon refleksinin tepki azalması ve güçlü uyaranlarla geçici restorasyon , J. Neurophysiol., 1966a, 29, 221-239.
Spencer WA, Thompson RF, Neilson DR, Jr. Yinelenen kutanöz afferent yaylımlarıyla aktive edilen yükselen ve refleks yollarının tepkisindeki değişiklikler, J. Neurophysiol., 1966b, 29, 240-252.
Spencer WA, Thompson RF, Neilson DR, Jr. Ventral kök elektrotonusunun azalması ve yinelenen kutanöz afferent volleyler tarafından üretilen hücre içi kaydedilmiş PSP'ler, J. Neurophysiol., 1966c, 29, 253-274.
Takeuchi A., Takeuchi N. Loligo'nun dev sinapsının sinaptik öncesi ve sonrası aksonlarındaki elektriksel değişiklikler , J. gen. Physiol., 1962, 45, 1181-1193.
Tanzi E. I fatti e le le induzioni nell' odierna istologia dei sistema nervoso, Riv. sp. Freniat. Med. bacak. Yabancılaşma, 1893, 19, 419-472.
Thompson RF, Spencer WA Alışkanlık: nöronal davranış substratlarının incelenmesi için bir model fenomen. Psikolog. Rev., 1966, 173, 16-43.
Thompson RF, Berger TW, Madden J. IV. Memeli CNS'deki hücresel öğrenme ve hafıza süreçleri, A. Rev. Neurosci., 1983, 6, 447-491.
Thorpe WH Hayvanlarda öğrenme ve içgüdü. Methuen, Londra, 1956.
Vidal JM Evcil kümes hayvanlarında evlat ve cinsel damgalama arasındaki ilişkiler: yaş ve sosyal deneyimin etkileri, Anim. Behav., 1980, 28, 880-891. Vinogradova OS Bilgi ve limbik sistem kaydı. İçinde: Nöral aktivite ve davranışta kısa vadeli değişiklikler. (editörler G. Horn ve R.A. Hinde), 1970, s. 95-140. Cambridge University Press, Cambridge.
Voronin LL Mikroelektrot koşullandırmanın nörofizyolojik mekanizmalarının incelenmesi. İçinde: Sovyet Araştırma Raporları (ed. CD Woody), 1976, Cilt. 2, s. 1-59. Beyin Bilgi Servisi, California Üniversitesi, Los Angeles.
Duvar PD Alışma ve omurilikte tetanik sonrası güçlenme. İçinde: Nöral aktivite ve davranışta kısa vadeli değişiklikler (eds. G. Horn ve R.A. Hinde), 1970, s. 181-210. Cambridge University Press, Cambridge.
Ağırlık FF, Erülkar SD Tekrarlayan postsinaptik aksiyon potansiyelleri ile sinaptik verici salınımının modülasyonu, Science, 1976, 193, 1023-1025.
Whitfield IC İşitme yolu. Arnold, Londra, 1967.
Wickelgren BG Habituation in spinal motorneurones, J. Neurophysiol., 1967a, 30, 1404-1423.
Wickelgren BG Spinal internöronların alışması, J. Neurophysiol., 1967b, 30, 1424-1438.
Woody CD Belleği, öğrenme ve daha yüksek işlev. Springer-Verlag, New York, 1982.
Young JZ Sefalopodların dev sinir liflerindeki kaynaşık nöronlar ve sinaptik temaslar, Phil. Trans. R. Soc M 1939, B229, 465-503.
BÖLÜM 2
KALIPLAMANIN BİYOKİMYASAL ETKİLERİ
, alışkanlık fenomenini hesaba katmıyordu . Bu görüşler temel olarak diğer öğrenme türleriyle ilişkili bilgilerin kaydedilmesine yönelik mekanizmalarla ilgiliydi. Bu nedenle, aşağıdaki tartışmada, "öğrenme" kavramı, alışma dışında çeşitli öğrenme biçimlerine atıfta bulunacaktır .
nöronlar arasındaki belirli bağlantıların aşamalı olarak zayıflamasına dayandığına dair güvenilir deneysel kanıtlar vardır . Buna karşılık, herhangi bir deneysel kanıt olmaksızın , diğer öğrenme türleriyle ilişkili belleğin, nöronlar arasındaki belirli bağlantıların güçlendirilmesini gerektirdiğine uzun zamandır inanılmaktadır. Amplifikasyonun, bir presinaptik nöronun uyarılması üzerine bir postsinaptik nöronda ortaya çıkan bir dürtü olasılığını arttırdığı ve böylece belirli bir sinir yolunda iletimi kolaylaştırdığı varsayılmıştır. Bu etki, uyarıcı sinapsların etkinliğini artırarak da elde edilebilir. Önleyici sinapsların etkinliğindeki benzer bir artış , presinaptik lifteki dürtü, postsinaptik nöronu daha güçlü bir şekilde inhibe ederse, tamamen farklı sonuçlara sahip olabilir . Young (Young, 1966) , öğrenmenin gereksiz yolları engelleyerek gerçekleştiğini öne sürdü. İnhibitör sinapsların etkinliğindeki bir artış, yolların bu fonksiyonel ayrışmasının altında yatan olabilir. Hem uyarıcı hem de inhibe edici sinapsların öğrenmeye dahil olması mümkündür. Bununla birlikte, çok çeşitli modellerin tek bir hücresel süreç temelinde - sinaptik bağlantıların güçlendirilmesi - inşa edilebilmesi önemlidir.
Merkezi sinir sistemindeki bilgi depolama yerleri biliniyorsa , sinaptik değişikliklerin hafıza organizasyonuna katılma olasılığı deneysel analize tabi tutulabilir. Bununla birlikte, çözülmesi çok zor olan tam da bu tür depoların anatomik lokalizasyonu sorunudur. Hücresel mekanizmaları analiz etmek için nerede bulunduklarını bilmek gerekir. Bu olmadan, bir biyoloğun elindeki en gelişmiş analitik yöntemler bile güçsüzdür. Örneğin, mikroelektrot kaydı ve elektron mikroskobu, nöronal sistemlerin olağandışı ince ayrıntılarını ortaya çıkarabilir, ancak bu yöntemleri kullanmadan önce, mikroelektrodun nereye yerleştirilmesi gerektiğini ve beynin 1 mm3'ten büyük olmayan hangi bölümünün izole edilmesi gerektiğini bilmek gerekir. histolojik kesitler ve müteakip analizler için. Daha fazla başarılı araştırma için, varsayılan değişikliklerin nerede meydana geldiğini belirlemek gerektiği açıktır. Alışmanın nöral mekanizmalarını anlama başarısının, sinir sisteminin nöronların ve sinapsların aktivitesinde kademeli değişikliklerin geliştiği bölümlerinin tam lokalizasyonuna bağlı olduğunu vurgulamak gereksiz olmayabilir .
Bellek süreçleri için kritik olan departmanları nasıl belirleyebilirsiniz? Sinaps verimliliğinde varsayılan değişiklikler çeşitli şekillerde ortaya çıkabilir. Örneğin, iki hücre arasındaki sinapsların sayısı artabilir veya postsinaptik aksonun terminal dalları artabilir. Her durumda, sonuç , presinaptik elyaftaki dürtünün, zarının daha geniş bir alanının geçirgenliğini değiştirerek, postsinaptik nöronun uyarılabilirliği üzerindeki daha güçlü bir etkisi olacaktır. Sinaptik etkinlikte bir artış, postsinaptik zar üzerindeki reseptör lokuslarının sayısını artırarak ve ayrıca bu zarın reseptör proteinlerinin şeklini veya konformasyonunu değiştirerek elde edilebilir. Bu makromoleküller , nöron yapısı ve işlevinin tüm yönleri için temel olduğundan , birçok sinaptik plastisite biçimi proteinleri içeriyor gibi görünmektedir . Artan protein tüketimi sentezini uyarabilir. Bu nedenle, bilgi depolamanın sinapsların verimliliğindeki artışa dayandığı hipotezi, etkilenen nöronlardaki sinaptik değişikliklerin protein sentezi oranını arttırdığı varsayımını içerir. Bu varsayım, belleğin nöral temelinin deneysel analizi için umut verici bir başlangıç noktası sağlar: Bir hayvan öğrendiğinde, iz depolamayla ilgili alanlarda protein sentezi hızı artmalıdır. Böyle bir değişim tanımlandıktan sonra , öğrenme süreciyle tam ilişkisi incelenebilir. Elbette böyle bir sentez artışının sadece depolama alanıyla sınırlı kalacağına inanmak saflık olur; en iyi ihtimalle, protein sentezinde bir artışın bulunacağı alanlar arasında , doğrudan * hafıza süreçleriyle ilgili olanların olacağı * umulabilir;
Protein metabolizması
Protein moleküllerinin yapı taşları amino asitlerdir. Dolayısıyla protein sentezi arttıkça amino asit alımı da artacaktır. Bu artış , yeni sentezlenen proteinlere dahil edilen radyoaktif izotoplarla işaretlenmiş amino asitler kullanılarak tespit edilebilir . Öğrenme prosedürünün bir sonucu olarak beynin herhangi bir bölgesinde protein sentezi hızı artarsa, o zaman işaretli amino asidin dahil edilmesi de artmalıdır. Buna göre, bu bölgedeki radyoaktivite eğitimli hayvanların beyinlerinde eğitimsiz kontrollerin beyinlerine göre daha yüksek olmalıdır.
Bu şekilde tepki veren beyin bölgelerini lokalize etmenin birkaç yöntemi vardır. En basit yöntemlerden biri, beyni birkaç bölüme ayırmak ve her birindeki radyoaktivite seviyesini ölçmektir. Eğitimli ve eğitimsiz hayvanların beyinlerinden alınan benzer örnekler için elde edilen değerler daha sonra karşılaştırılabilir. Beynin toplam metabolik aktivitesinin dağılımının oldukça doğrudan bir değerlendirmesine izin veren bu yöntemin avantajı, oldukça basit olmasıdır. Bir hayvana, çok çeşitli proteinlere dahil edilmiş bir radyoaktif amino asit enjekte edilir. Böyle bir amino asit lizindir. Lizin, radyoaktif bir hidrojen izotopu olan trityum (3H) ile etiketlenebilir . Amino asidin enjeksiyonundan kısa bir süre sonra hayvan eğitilebilir. Öğrenme süreci sırasında, dolaşımdaki 3H - lizinin bir kısmı yeni sentezlenen proteine dahil edilecektir. Katılan habercinin miktarını belirlemek için hayvan kurban edilir ve beyin bölgeleri hızla izole edilir. Analizin başlangıç aşaması, numunenin homojenleştirilmesi ve homojenat kısmındaki protein içeriğinin belirlenmesidir. Diğer kısma trikloroasetik asit eklenir . Asit, bazıları 3 H-lizin içeren proteinleri çökeltir. Dahil olmayan 3 H-lizin solüsyonda kalır (Şekil 2.1). Çökelti veya asitte çözünmeyen fraksiyon süpernatandan ayrılır ve içindeki radyoaktivite seviyesi ölçülür. Radyoaktivite, 1 mg protein başına 1 dakikada bozunma sayısı olarak ifade edilir (spesifik aktivite); bu, etiketli bir amino asidin bir proteine dahil olma seviyesinin bir ölçüsünü verir.
Şaşırtıcı olmayan bir şekilde, bu yöntemin sınırlamaları vardır. İlk olarak, onun yardımıyla sadece kaba anatomik lokalizasyon gün ışığına çıkar. İkincisi, protein sentezi ile etiketlenmiş öncül birleşme arasındaki ilişki o kadar basit değildir ve bu tür çalışmaların sonuçları büyük bir dikkatle yorumlanmalıdır (bakınız Hom ve diğerleri, 1973). Her şeye rağmen,
civciv damgalamayla ilgili ilk biyokimyasal çalışmamız için etiketli progenitör yöntemini seçtik (Bateson ve diğerleri, 1972). Bu çalışmanın, beynin herhangi bir bölümünün seçici aktivasyonunun bu öğrenme biçimiyle gerçekleşip gerçekleşmediğine dair ilk belirtileri sağlaması gerekiyordu .

Çalışmamızda yumurtadan çıkan civcivler, damgalamanın kolay olduğu bir gelişim aşamasındaki bir nesne ile sunulana kadar karanlıkta tutuldu. Bu deneysel kuşlarda, radyoaktif lizinin beynin üç bölgesine dahil edilmesi ölçüldü. Tavukların iki kontrol grubu için de karşılık gelen rakamlar elde edildi. Bu gruplardan biri ışıkta tutuldu, ancak damgalayıcı bir uyarana maruz bırakılmadı; ikinci kontrol grubunun tavukları karanlıkta tutuldu. Deney grubuna damgalama ve yabancı nesneler arasında bir seçim vererek öğrenme derecesinin doğrudan bir ölçümü yoktu . Aynı damgalama uyaranına maruz kalan tavukların onu yeni bir nesneye tercih ettiği önceki deneylere dayanarak, öğrenmenin gerçekleştiği kabul edildi ( Bateson ve Wainwright, 1972).
Kullanılan damgalayıcı uyaran, Şekil 2'de gösterilene benzer bir formdaydı. 2.2, A. Kutunun iki büyük dairesi turuncuya boyanmıştır. 1 dakikada 85 devir frekansına sahip Kutu Raschalas. Yaklaşık 4* sonra



Yumurtadan çıktıktan 17 saat sonra, deney grubundaki civcivler özel bir alanda ayrı bölmelere yerleştirildi (Şekil 2.3). Kalemlerin zemini ve üç duvarı opaktı. Platformun merkezine bakan dördüncü duvar şeffaftı ve civciv bunun içinden yanıp sönen turuncu ışığı görebiliyordu. Kalemlikler yukarıdan açıktı ve tüm saha 60 W'lık bir ampulle aydınlatılıyordu. Işıkla kaldırılan kontrol grubundaki tavuklar benzer bölmelerdeydi, ancak titreşen ışığa bakan duvar opaktı. Karanlıkta yetiştirilen üçüncü grup civcivler ise koyu renkli yoğun malzemeyle kaplı kümeslerde tutuldu. Böylece, tüm tavuk grupları, hafif bir uyaranı döndüren bir motorun gürültüsüne maruz bırakıldı , iki grup üstten aydınlatma aldı (deneysel ve "hafif" gruplar), ancak yalnızca bir deney grubu turuncu kutuyu
görebildi .

Resim: 2.3. Deney civcivlerinin ( e.c. ) dönen flaş ışığına (s.c.) ve "ışık kontrol" (s.c.) grubundaki civcivlerin üstten aydınlatmaya maruz bırakıldığı kümes planı. Deney civcivleri, şeffaf panelden (kesikli çizgi) yanıp sönen ışığı görebildi . Diğer tüm duvarlar opaktı (düz çizgiler). Platform yukarıdan açıktı ve ağıllarla aynı şekilde tepeden bir elektrik ışığıyla aydınlatılıyordu. (Bateson ve diğerleri, 1972.)
Resim: 2.4. Ördek yavrularının tanıdık bir Ördek ile aynı türden alışılmadık bir yem arasında seçim yapabilecekleri bir testin sonuçları. Takip etme ve damgalama tepkisinin oluşumunun yaşa ve öğrenmeye bağlılığı gösterilmiştir. Sütunların altındaki göstergeler, inkübasyonun başlangıcından eğitimin başlangıcına kadar olan günleri ve saatleri gösterir. Ördek yavrularının, her iki ördeğe eşit veya yalnızca bilinmeyen ördeğe yaklaşmaları durumunda aşağıdaki tepkiyi (daireler) verdiğine inanılıyordu. Çoğunlukla tanıdık bir ördeği (eğik trich) takip eden bir ördek yavrusu damgalanmış olarak kabul edildi. Sütunun her bir bölümünün yüksekliği, belirtilen davranışlara sahip ördek yavrusu sayısını gösterir; örneğin, yüksekliği olup olmadığı ° ve LbIK ' ile rastgele düzenlenmiş kısa dalgalı Van Yami ' coo J BeTCTB y eT reaksiyon göstermeyen ördek yavrusu sayısı
İki kontrol grubuna ek olarak, deneyde bazı ek kontrol prosedürleri kullanıldı. Bu, üç grup tavuğun beyin dokusunda tespit edilebilecek herhangi bir biyokimyasal farklılığın davranışsal bağıntılarını açıklığa kavuşturmak için yapıldı . Tanıtılan kontrol prosedürü Gottlieb'in (Gottlieb, 1961) çalışmasına dayanıyordu . Ördek yavrusu yeşilbaş gösterdi (Anas platyrhynchos) doldurulmuş ördek. Daha sonra her ördek yavrusu aynı ve başka bir doldurulmuş hayvan arasında seçim yapmasına izin verildi. Gottlieb, eğitim sırasında sunulan doldurulmuş bir hayvanı seçici olarak takip etme vakalarının sıklığının, kuluçka başlangıcından itibaren sayılan ördek yavrularının yaşına bağlı olduğunu buldu (Şekil 2.4). Evcil tavukların tavuklarında da benzer bir fenomen gözlenirse, o zaman iki yaş grubu seçilebilir ve bunlardan birinde ışıklı kutuya başarılı baskı yapma şansı elde edilebilir ve aynı uyarana maruz kalan diğerinde bu olasılık azaltılabilir. Beyindeki herhangi bir biyokimyasal kayma, damgalama süreciyle ilişkiliyse, bu grupların birincisinde bulunmalı ve ikincisinde bulunmamalıdır. Bu nedenle civcivlerin yarısı ana çıkıştan 6-9 saat önce çıkanlardan, yarısı ise pikten 6-9 saat sonra çıkanlardan alınmıştır. Bu iki tavuk grubu sırasıyla "erken" ve "geç" olarak belirlendi.
Deneyin seyri her iki grup için de aynıydı. Deneysel tavuklar, onlara titreyen bir ışık gösterilerek eğitildi. "Hafif" kontrol grubu bu uyarıcıyı görmedi, ancak kalemleri, merkezde asılı duran bir elektrik ışığıyla yukarıdan aydınlatıldı. Deneyin toplam süresi her iki grup için 115 dakika idi. Bu süre zarfında, "karanlık" kontrol grubu kapalı muhafazalarda tutuldu. Bu sürenin bitiminden 40 dakika önce her tavuğa kalp bölgesine 20 μCi 3H-lizin hızla enjekte edildi. Deneyin sonunda (115 dk sonra) tüm civcivler 40 dk kalacakları karanlık bir yere konuldu. Daha sonra kısa (2 dakika) bir teste tabi tutuldular. Bu durumda, her bir tavuk, bir ucunda deney grubuna maruz kalanla aynı olan, titreşen turuncu bir ışığın olduğu bir koridora yerleştirildi. Aynı zamanda, her bir tavuğun çıkardığı kaygı ve zevki gösteren seslerin sayısı kaydedildi (Collias ve Joos, 1953; Andrew, 1964).
Kesimden sonra, tavukların beyinleri hızla çıkarıldı ve üç parçaya bölündü (Şekil 2.5). Ön beyin yarım küreleri, yarım kürelerin ön kutuplarından arka kutuplarına doğru uzanan eğik bir kesi ile ikiye bölünmüştür. Bu kesit planının seçilmesinin nedeni , kesitten dorsal olarak uzanan beynin ön kısmının tüm komayı içermesidir.

B
Ryas. 2.5. Tavuk beyni (yandan görünüm). A. Radyoaktif lizinin proteinlere dahil edilmesi çalışmasında kullanılan alanlar : önbeynin dorsal kısmı ("çatı ") ventral kısımdan ("taban") kalın kesikli bir çizgiyle gösterilen bir kesikle ayrılmıştır; orta beyin optik çatıyı içerir. Beynin kullanılmayan kısımları ince kesikli çizgilerle gösterilmiştir. B. Ön beyindeki belli başlı bölgelerin ana hatları çizilir ve orta hatta paralel ama biraz yanal uzanan bir kesikle bölünür. 1 - ventrikül; 2 - ek hiperstriatum; 3 - orta hiperstriatum; 4 - dorsal hiperstriatum. Bu yapıların üçü birlikte görsel şehveti oluşturur; 5 - neostriatum; 6 - yükselen paleostriatum; 7, ilkel paleostriatum.
(görsel Wulst) ilişkili hiperstriatumun bileşenleri . Orta beyin ön beyinden ve arka beyinden optik çatının büyük loblarından anterior ve posterior olarak iki dikey kesi ile ayrıldı. Arka beyin ve beyincik atılırken, retinadan ana girdiyi alan optik çatı da dahil olmak üzere orta beyin korunur. Beyin parçaları, katı karbon dioksit içeren kaplarda ayrı ayrı donduruldu ve -20°C'de saklandı. Her kap bir kodla etiketlendi, böylece verilen beyin dokusu örneğinin ait olduğu tavuğun hangi davranışsal deneyime sahip olduğu bilinmeden biyokimyasal analiz yapıldı . Spesifik aktivite, asitte çözünmeyen bir çökeltide belirlendi. Ortaya çıkan değer, radyoaktif öncünün ne kadarının proteine dahil edildiğini gösterdi.
Deneyler birkaç hafta boyunca gerçekleştirildi, böylece tavuklar birkaç parti yumurtadan alındı. Aralarındaki farkları hariç tutmak için, asitte çözünmeyen fraksiyonun spesifik aktivitesi, belirli bir tavuk partisinin "karanlık" kontrolünden alınan tüm beyin örneklerinin ortalama değerine bölündü. Bu şekilde standardize edilen sonuçlar , "karanlık " kontrol grubu için ortalama değerin yüzdesi olarak ifade edildi.
Deneyin sonundaki kısa testte, erken gruptaki civcivler , geç gruptaki civcivlere göre önemli ölçüde daha az imdat çağrısı ve daha fazla zevk çağrısı yaptı. Geç gruptaki civcivlerdeki yüksek kaygı çağrıları, deney sırasında onların damgalama için en uygun gelişim aşamasını çoktan geçtiklerini gösterdi.
standardize edilmiş spesifik aktivitesinin göstergeleri, şekil 2'de gösterilmektedir. 2.6. Damgalayıcı uyaranın etkisinin etkisi , yalnızca bir grup tavukta - erken grupta - ve beynin yalnızca bir bölgesinde - ön beynin çatısında bulundu. Bu alanda, etiketin deney grubuna dahil edilmesi, "karanlık" kontrole göre önemli ölçüde daha yüksekti. "Hafif" kontrol grubundaki standartlaştırılmış spesifik aktivite , diğer iki grubun göstergeleri arasında bir ara pozisyon işgal etti (Şekil 2.6, L).
Bu deneylerde elde edilen diğer iki sonuç özellikle ilgi çekicidir. Titreşen ışığa maruz kalan tavuklarda ve daha az ölçüde "hafif" kontrol grubunda, impuls patlamaları retinadan optik çatıya ve oradan, muhtemelen beyin sapı ve ön beyine doğru gelmelidir (bkz. Bölüm A.3.1). ve Şekil S.5). Tüm deney boyunca karanlıkta tutulan tavukların retinalarından gelen deşarjların , deney tavuklarıyla aynı yoğunluğa ulaşması pek mümkün değildir. Bununla birlikte, radyoaktif lizinin orta beyne dahil edilmesi , erken ve geç grupların yanı sıra üç grubun hepsinde pratik olarak aynıydı. Bu nedenle, uyarılmış görsel dürtü aktivitesi seviyesindeki farazi farklılıklar , her biri geniş bir projeksiyon görsel alanı içeren, analiz için alınan beyin parçalarında habercinin dahil edilmesindeki farklılıklarla ilişkili değildi . Bununla ilgili ikinci nokta, ön beyin çatısıyla ilgilidir. Hem erken hem de geç deneysel tavuk grupları titreşen ışığa maruz bırakıldı. Bununla birlikte, yalnızca erken grupta, radyoaktif lizinin dahil edilmesinde önemli bir artışla ilişkili deneyim kazanımı vardı . Bu nedenle, orta beyin durumunda olduğu gibi, görsel deneyim bir taco gibidir.

N 15 15 16 15 15 16 15 15 16
tavuk sayısı
Resim: 2.6. Deneysel (siyah çubuklar), "açık" kontrol (ızgara ile kaplı çubuklar) ve "koyu " kontrol (gölgeli çubuklar) gruplarında üç beyin bölgesindeki 3H -lizin içeren proteinler için standart spesifik aktivite değerleri . A. Erken çıkan civcivler. Deney grubundaki ön beyin çatısı için göstergeler, "karanlık" kontrol grubundan daha yüksektir (P<0.05). Gruplar arasında başka anlamlı fark yoktur. B. Geç yumurtadan çıkan civcivler . Beynin farklı bölümleri için göstergeler tüm gruplarda önemli ölçüde farklılık göstermez. Bu ve sonraki çizelgelerde çubukların altındaki N değerleri, gruptaki civciv sayısıdır. (Bateson ve diğerleri, 1972.)
işaretli amino asidin dahil edilmesinde nispi bir artış için yeterli değildir.
Erken grup tavuklarda radyoaktif lizinin ön beyin çatısına artan şekilde dahil edilmesinin ve geç grupta bu etkinin olmamasının , imprinting prosedürünün spesifik bir etkisinin kanıtı olduğuna dair cazip bir öneri ortaya çıkıyor. Ancak bu varsayım, yalnızca yukarıda açıklanan deneylere dayanarak geçerli kabul edilemez. Örneğin, yumurtadan erken çıkan civcivler geç çıkan civcivlerden daha aktif olabilir (Davies ve Raupe, 1982). Bu durumda, erken civcivlerde gözlemlenen spesifik biyokimyasal değişiklikler, yüksek düzeyde lokomotor aktiviteden kaynaklanıyor olabilir ve damgalama ile ilgisi olmayabilir. Bu nedenle, biyokimyasal etkilerin davranışsal bağıntılarının yorumlanmasında bir belirsizlik vardır. Bu belirsizliğin nasıl önleneceği Bölüm 1'de açıklanacaktır. 3. Bu çalışmanın sonuçlarının temkinli yorumlanması için başka nedenler de vardır; örneğin, görsel stimülasyon olmadığında bile etiketli lizin birleşmesi hızlıdır . Bu gerçek, haberci dahil etme oranının , damgalama prosedürünün neden olduğu protein sentezindeki olası kaymaların nispeten duyarsız bir göstergesi olduğunu göstermektedir . Biyokimyasal kaymaların daha hassas bir göstergesinin gerekli olduğu açıktır.
RNA değişimi
Ribonükleik asit molekülleri, proteinlerin sentezi için gereklidir. Amino asitler, ribozomlar üzerinde birleşmek için transfer RNA ile birleşir. Ribozomlar, farklı proteinlerin yapısını belirleyen birkaç RNA molekülü ile ilişkili büyük protein kompleksleridir. Her biri bir amino asit taşıyan transfer RNA molekülleri, bir ribozom üzerinde toplanır ve haberci (matriks) RNA'dan genetik bilgiyi okur. Messenger RNA, protein makromoleküllerindeki amino asitlerin dizisini ve sayısını belirler. RNA'nın protein sentezindeki bu merkezi rolü göz önüne alındığında, bu sentezin yoğunluğundaki değişikliklere RNA metabolizmasındaki değişikliklerin eşlik edeceği varsayılabilir . Radyoaktif lisinin asitte çözünmeyen maddelere dahil edilmesi protein sentezindeki değişiklikleri yansıtıyorsa, bu tür kaymaların damgalama nedeniyle ortaya çıkması olasıdır (Bölüm 2.1). Bu varsayımlar doğrultusunda dikkatimizi RNA'ya çevirdik.
belirli sayıda ribonükleotidin oluşturduğu bir makromoleküldür . Her nükleotid bir şeker molekülü (riboz), bir fosfat grubu ve dört bazdan (adenin, guanin, sitozin veya urasil) birini içerir. İlk üç baz DNA'da da bulunuyorsa, urasil DNA'da asla bulunmaz. Ön çalışmalar, seyirci yokluğunda
Grup N

Resim: 2.7. Radyoaktif urasilin haberci olarak kullanıldığı bir deneyin şeması. Tüm civcivler 20 μCi 3H-urasil uygulamasından sonra ve son flaş ışığı testine (beyaz bantlar) kadar karanlık bir inkübatörde tutuldu . Deney grubunun yukarıdan aydınlatmaya ve titreşen ışığa maruz kaldığı süre siyah çubuklarla gösterilmiştir. Üst aydınlatmanın "hafif" kontrol grubu üzerindeki etki süresi, taranmış çizgilerle gösterilir. 76 dakika uyarılan deney ve "hafif" gruplar, enjeksiyondan 37 dakika sonra ağıllara yerleştirildi. 38 dakika uyarılan civcivler enjeksiyondan 56 dakika sonra kümese yerleştirildi. Enjeksiyondan tıkamaya kadar geçen toplam süre 150 dakikaydı. N, her gruptaki tavuk sayısıdır.
işaretli urasil, stimülasyondan sonra çok daha yavaş açılır ve bu nedenle, etiketli lizinden daha metabolizma seviyesinin daha hassas bir göstergesi olabilir. Bu temelde, şeması Şekil 1'de gösterilen bir sonraki deneyde. 2.7, öncü olarak urasil kullanıldı.
Deney grubu, "aydınlık" kontrol ve "karanlık" kontrol gruplarının yanı sıra, ilk deney serisindekiyle aynı koşullara tabi tutuldu. Civcivler erkenci ve geçci gruplara ayrılmamış, sadece kuluçka döneminin başında ve ortasında çıkanlar alınmıştır. Deney ve “hafif” gruplar 38 veya 76 dakika boyunca uygun koşullara maruz bırakıldı . Bu iki dönem, 30 dakika boyunca damgalama uyaranına maruz kalan civcivlerin (Şekil 2.2, A ve 2.3) bunu tercih etmemesi, yani damgalama yapmamaları nedeniyle seçildi. Buna karşılık, 60 dakika boyunca uyarılan tavuklar bir tercih geliştirdi, yani damgalandı (Bateson, 1974). Bu nedenle, 38 dakika sonra haberci birleştirme kaymaları olmazsa, 76 dakika sonra ortaya çıkabilirler. Deneyin sonunda beynin bölgeleri izole edildi ve şifrelendi. Daha sonra, beyin homojenatının asitte çözünmeyen fraksiyonunda RNA saptandığından, 3H-lizin ile yapılan deneylerde kullanılana benzer bir biyokimyasal analiz gerçekleştirildi .

Etiketli urasil'in asitte çözünmeyen fraksiyona dahil edilmesinin , yalnızca bir grup tavukta ve beynin yalnızca bir bölgesinde "karanlık" kontrole kıyasla arttığı bulundu . 76 dakika süreyle damgalama uyaranına maruz kalan tavuklarda bir artış bulundu (Şekil 2.8). Bu grupla ilgili olarak, damgalamayı geliştirdiklerini varsaymak güvenliydi . Böyle bir artış yalnızca, önceki deney serilerinde (Bölüm 2.1) değişikliklerin de bulunduğu bir alan olan önbeynin çatısında gözlemlendi .
Anlamlandırmada bazı zorluklar
Açıklanan iki dizi deneyin olumlu sonuçları arasında, biyokimyasal değişikliklerin görsel bir nesnenin etkisiyle ilişkili olduğunu ve bu değişikliklerin beynin belirli bir bölgesinde meydana geldiğini gösteren sonucu içermelidir . Davranış çalışması için bu verilerin önemini düşünmeden önce , bunların biyokimyasal sonuçları analiz edilmelidir. Her iki deney de birbirini doğrulamaktadır, ancak bunların yorumlanmasında , etiketli bir habercinin dahil edilmesinin analiziyle elde edilen tüm sonuçlarda ortak olan bazı zorluklar vardır . Artan protein sentezi, radyoaktif amino asidin asitte çözünmeyen maddelere daha fazla dahil olmasına neden olsa da, bundan, protein sentezinin fiilen arttığı sonucu çıkmayabilir . En iyi ihtimalle, haberci birleştirme, saf sentezden ziyade döviz kurlarının göstergesidir. Diğer bir zorluk, makromoleküllerin öncüllerinin hücre içinde yer almasından kaynaklanmaktadır. Bu nedenle , radyoaktif haberci nöronun içine nüfuz etmelidir. İşaretli öncü, kan dolaşımındaki hücrelere iletilir ve belirli bir alana kan temini, farklı fizyolojik koşullar altında büyük ölçüde değişir. Buna göre , etiketli habercinin hücre yüzeyi ile temas derecesi ve muhtemelen dahil edilebilecek haberci miktarı değişecektir . Hücre zarı boyunca haberci alım ve penetrasyon mekanizmalarının çeşitliliği ve ayrıca etiketlenmemiş öncüllerin havuzlarındaki farklılıklar, yorumlamayı daha da zorlaştırır.
Bu zorlukların bazıları, in vitro tahlil yöntemleri kullanılarak aşılabilir . Bu yöntem, in vivo dahil etme çalışmalarını çok zorlaştıran haberci havuzunun boyutundaki dalgalanmaları ortadan kaldırma avantajına sahiptir. Bu yöntemi kullanarak, Haywood ve diğerleri ( 1970) , damgalamanın ön beyin çatısında artan RNA sentezi ile ilişkili olduğuna dair kanıt elde ettiler; beynin tabanında veya orta beyinde RNA sentezi artmaz.
Genel olarak, biyokimyasal veriler aşağıdaki değişim zincirine karşılık gelir: bireysel deneyim -"- ön beyin çatısındaki nöronları etkileyen nabız aktivitesi -> haberci RNA oluşumu ile DNA transkripsiyonu -> protein sentezi.
Protein sentezinde artışa yol açan süreçler, kadın cinsiyet hormonu östrojen gibi belirli steroid hormonları tarafından da etkinleştirilebilir. Bununla birlikte, damgalama durumunda belirtilen olaylar zincirinin temelde mantıksal bir yapı olduğunu vurgulamak önemlidir (Horn ve diğerleri, 1973). Kuşkusuz, titreyen ışığa 30 dakika maruz kaldıktan sonra RNA polimerazın aktivitesi arttı , 3H-urasil'in RNA'ya dahil edilmesi 76 dakika stimülasyondan sonra meydana geldi ve 3H-lizinin proteine dahil edilmesindeki değişiklikler 115 dakikalık eğitimden sonra gözlendi . . Bununla birlikte, bu dizi doğru olmayabilir , ancak bariz olabilir: RNA sentezindeki zaman içindeki değişikliklerin dinamikleri , nesnenin maruz kalma süresi değiştirilerek analiz edildi, ancak benzer bir analiz, protein metabolizmasını incelerken yapılmadı . Önerilen biyokimyasal modelin doğru olup olmadığı, biyokimyasal kaymalar ve baskılama arasındaki ilişki sorgulanabilir. Bu şüphelerin doğası ve onlardan kurtulmaya çalıştığımız yollar bir sonraki bölümde anlatılacaktır.
Özet
Birçok öğrenme biçimi, bir yanıt geliştirmeyi veya onu belirli deneyimlerle ilişkilendirmeyi içerir. Bu "kademeli" öğrenme ve depolama biçimlerinin altında yatan nöral değişiklikler, genellikle nöronlar arasındaki sinaptik bağlantıların güçlendirilmesi olarak tanımlanır. Bu hipotezin olası, ancak zorunlu olmayan bir sonucu vardır, bu da beynin eser miktarda depolandığı bölümlerinde protein sentezinin arttırılması gerektiğidir. Tavuklarda damgalama üzerine yapılan bir çalışmadan elde edilen veriler, açıkça tanımlanmış bir görsel nesneye maruz kalmanın, bir radyoaktif amino asidin bir proteine ve bir radyoaktif urasilin RNA'ya dahil edilmesindeki artışla ilişkili olduğunu göstermiştir. Bu biyokimyasal sonucun yorumlanmasındaki bazı zorluklar tartışılmıştır.
EDEBİYAT
Andrew RJ Civcivlerde Seslendirme ve “uyarıcı” kavramının karşıtlığı, Anim. Davranış, 1964, 12, 64-76.
Bateson PPG Civcivlerde eğitim süresi, karşılaştırma fırsatları ve damgalama, J. comp. fizik. Psychol., 1974, 86, 586-589.
Bateson PPG, Wainwright AAP Daha önce ışığa maruz kalmanın evcil civcivlerde baskı süreci üzerindeki etkileri, Behavior, 1972, 42, 279-290.
Bateson PPG, Horn G., Rose S. PR Erken deneyimlerin öncülerin civciv beyninde RNA ve proteine bölgesel olarak dahil edilmesi üzerindeki etkileri, Brain Res, 1972, 39, 449-465.
Cottias N., Joos M. Evcil kümes hayvanlarının ses sinyallerinin spektrografik analizi, Behavior, 1953, 5, 175-187.
Davies DC, Payne J. Yumurtadan çıkma sırasında civciv cinsiyet oranlarındaki değişim, Anim. Davranış, 1982, 30, 931-932.
Gottlieb G. Damgalamada kritik dönemin belirlenmesi için bir temel olarak gelişimsel yaş, J. comp. fizik. Psychol, 1961, 54, 422-427.
Haywood J., Rose SPR, Bateson PPG Civciv beyninde RNA polimeraz aktivitesi üzerinde bir damgalama tercihinin etkisi, Nature, Lond, 1970, 228, 373-374.
Horn G., Rose SPR, Bateson PPG Merkezi sinir sisteminde deneyim ve plastisite, Science, '1973, 181, 506-514.
Young JZ Beynin hafıza sistemi. Oxford University Press, Londra, 1966.
BÖLÜM 3
BİYOKİMYASAL DEĞİŞİKLİKLER ÖĞRENMENİN BİR TESPİTİ MİDİR?
Biyokimyasal değişiklikler ve damgalamanın ilişkili olduğu gerçeği , öğrenmenin nöronlar arasındaki bağlantılardaki değişikliklere dayandığı fikriyle tutarlıdır . Bununla birlikte, biyokimyasal değişimler çeşitli nedenlerle ortaya çıkabilir ve çeşitli süreçlerle ilişkilendirilebilir, öğrenme bunlardan sadece biridir. Titreşen ışığa maruz kalan civcivler , karanlıkta kontrollere göre daha fazla hareket edebilir, daha uyanık, ajite ve hatta daha stresli olabilir. Ek olarak, bu kontrol civcivlerinin, kursiyerlerin aksine, görsel deneyimleri yoktur veya çok azdır. Bu iki grup tavukta ön beyin çatısında gözlenen biyokimyasal farklılıklardan bu faktörlerin herhangi biri veya bunların bir kombinasyonu sorumlu olabilir . Örneğin beyindeki protein ve RNA metabolizması stresin etkisi altında değişir (Bryan ve diğerleri, 1967; Jakoubek ve diğerleri, 1970). Stresli bir durum adrenalin, büyüme hormonu, kortikotropin ve kortizol salgılanmasını etkileyebilir. Bu hormonlar sırayla, amino asitlerin proteinlere dahil olma oranını ve RNA sentezi oranını etkileyebilirler (Wagle, 1963; Jakoubek ve diğerleri, 1972). Bu nedenle, nöronal organizasyondaki öğrenme ile ilgili değişiklikler, karanlıkta barındırılan ve eğitilmiş tavuklar arasındaki farklılıkları açıklayabilirken , bu etkiyi yalnızca veya hatta kısmen öğrenme süreçlerinin etkisine atfetmek tamamen yanıltıcı olabilir.
Yukarıdakilerin hepsi apaçık görünebilir, ancak deneyimler, öğrenmenin gerçek etkisi olan sinirsel değişiklikleri, olmayan yan etkilerden ayırmanın son derece zor olduğunu göstermiştir . Öğrenme sürecinin bu nöronal sonuçlarını ayırt etmedeki ilerleme çok yavaş olmuştur. Örneğin, eğitimli hayvanlar ile eğitimsiz bir kontrol grubu arasındaki lokomotor aktivitedeki farklılıkları tespit etmek için tasarlanmış bir deneyde, her grup tarafından alınan duyusal uyarının miktarı ve derecesindeki farklılıklar dikkate alınmayabilir. Duyusal stimülasyondaki farklılıkları kontrol ederken, iki hayvan grubunun motivasyonel durumu üzerindeki kontrol gözden kaçabilir, vs. Tek başına hiçbir deney , öğrenme sürecinin tüm yan etkilerini kontrol edemez. Ancak bu, belirli bir yan etkinin beyin durumunda gözlenen değişikliği tamamen belirleme olasılığını azaltmak için bir dizi deneyde yapılabilir . Buna göre , bu öğrenme şekli için damgalamayla (Bölüm 2) ilişkili gözlemlenen biyokimyasal değişikliklerin özgüllüğünü belirlemek için bir dizi kontrol prosedürü geliştirdik .
beyni bölmek
ve stresin etkisi altında değişebilmektedir . Flaş ışığına maruz kalan tavukların ön beyin çatı hücrelerine işaretli urasil inkorsiyonundaki artış bu faktörlere bağlıysa, o zaman bu tür bir artış , hariç tutulursa gerçekleşmemelidir. Örneğin, karanlıktaki civcivler ile damgalayıcı bir uyarana maruz kalan civcivler arasındaki biyokimyasal farklılıkların , yalnızca hormonal düzeylerde stres kaynaklı bir farklılığın sonucu olduğunu varsayalım . Ayrıca , bir tavuğun beynin bir tarafının diğer tarafını etkilemeden "eğitilebileceğini" varsayalım . Böyle bir beynin "öğrenilmiş" tarafı, "öğrenilmemiş" tarafa kıyasla hormonların etkisinden dolayı daha yüksek düzeyde öncü birleşme ile karakterize edilecek mi? Muhtemelen hayır, çünkü hormonlar kan dolaşımına salınır ve beynin her iki tarafı da bunlara maruz kalır. Bu nedenle, "öğrenilmiş" tarafta dahil olma daha yüksekse, o zaman damgalayıcı uyaranın etkisinin bu biyokimyasal etkisi, kandaki hormon seviyesindeki bir değişiklik temelinde kolayca açıklanamaz. Bir "eğitimli" beyin yarım küresine sahip tavukların , eğitimin spesifik olmayan etkilerini tespit etmek için yararlı bir kontrol işlevi görebileceği açıktır.
Myers, Sperry ve Henson'ın (Myers, Sperry, 1953; Myers, 1956; Myers, Henson, 1960) ilk çalışmalarından sonra , yazarların öğrenmenin en azından sınırlanabileceğini belirten verilerden alıntı yaptıkları çok sayıda çalışma ortaya çıktı. tezahürlerinde, beynin bir tarafı. Kuşlarda tek taraflı öğrenme için gerekli prosedür Cuenod ve Zeier (Cuenod ve Zeier, 1967) tarafından geliştirilmiş ve daha sonra Meier (Meier, 1971) tarafından güvercinler (Columba IIIa) üzerindeki çalışmalarında kullanılmıştır. Bu kuşlarda, optik kiazma tamamlanmıştır (Cowan ve diğerleri, 1961), böylece sol gözden gelen sinirler beyin sapının sağ tarafına geçer.

Resim: 3.1. Bozulmamış (L) ve komisyonlanmış (B) güvercinlerde şekil ayrımı için öğrenme eğrileri. Tanınabilir rakamlar eğrilerin üzerinde gösterilmiştir. Güvercinler, tek gözü (ilk göz) kullanarak figürleri ayırt etmeyi öğrendi; ikincisi o sırada kapalıydı. Kritere (kesikli çizgi) ulaşıldığında, başlangıçta kapalı olan (ikinci) göz kullanılarak eğitime devam edildi. Bozulmamış güvercinler (A) sadece diğer gözü kullandığında, farklılaşma yanıtı, başarılı bir öğrenme transferini gösteren kritere hemen ulaştı. Dorsal supraoptik komissürün (B) transeksiyonunda, sadece ikinci gözün kullanıldığı eğitim süresi neredeyse ilk göz kadar uzundu. Benzer sonuçlar 46 kontrol güvercininde ve dorsal supraoptik komisyonu kesilmiş 9 güvercinde elde edildi.
(Şekil A.5). Dorsal supraoptik komissürün bir parçası olarak orta hattı geçen bir bileşen (Şekil P.5, B) dışında, ön beyine yapılan diğer çıkıntıların çoğu da çapraz olarak kalır (Şekil P.5, E ). Ön beyinde iki ana görsel projeksiyon alanı vardır. Bunlardan ektostriatum yalnızca kontralateral gözden girdi alır. Diğer alan, görsel Wulst , her iki gözden de sinyal alır. Dorsal supraoptik komissürün kesilmesinden sonra, hemisfer sadece kontralateral gözden girdi alacaktır.
Meyer (Meier, 1971), güvercinlerde öğrenme sürecini incelerken anatomik organizasyondaki bu avantajdan yararlandı. Önce kuşun bir gözünü kapattı ve ardından güvercini açık (“birinci *”) gözü kullanarak iki uyaran konfigürasyonunu ayırt etmesi için eğitti (Şekil 3.1). Kuşun gıda takviyesi alabilmesi için bir uyarıcıyı (pozitif uyarıcı) gagalaması gerekiyordu. Birbirini takip eden iki deneyin ilk 20 denemesinde %90 doğru cevap verme kriterine kadar birkaç gün boyunca eğitim gerçekleştirildi . Kritere ulaştıktan sonra kuşlar test edildi ve başlangıçta kapalı olan (“ikinci”) gözü kullanmaları gerekiyordu. Tüm kuşlar, "ilk" gözü kullanırken gerekli olandan daha az sayıda denemeyle kritere ulaştı (Şekil 3.1, R). Böylece bir “öğrenme aktarımı” gözlemlenmiştir. Meyer daha sonra bu deneyi tekrarladı, ancak dorsal supraoptik komissürün kesilmiş olduğu kuşları kullandı (Şekil 3.1b). "Birinci" gözü kullanarak kritere ulaşmak için gereken deneme sayısı, bozulmamış kontrol kuşlarındakiyle aynıydı. Bununla birlikte, eğitim sırasında kapalı "ikinci" göz kullanılarak kesik komissürlü kuşları test ederken, becerinin kalıcı olduğuna dair hiçbir işaret bulunmadı. Bu sonuçlar , bozulmamış bir güvercindeki figürleri tek gözle ayırt etmeyi öğrenirken, öğrenmenin diğer gözün temsiline aktarıldığını göstermektedir. Bu transfer , dorsal supraoptik komissürün kesilmesinden sonra gerçekleşmez.
Örneğin, sol retinaya etki eden görsel bir görüntü tarafından sol optik sinirde uyarılan impuls aktivitesi, önbeynin her iki yarımküresine de ulaşır (Perisic ve diğerleri, 1971; Brown, Hoot, 1979). Bu nedenle, sağlam bir güvercinin sağ gözü kullanılarak yapılan monoküler testinde , kuşun şu kriteri elde etmesi için gereken önemli ölçüde daha az sayıda örneğe ihtiyacı vardır : sağ göz her iki yarım küreye bağlıdır ve bu bağlantılar, içlerinde depolanan bilgilere erişim sağlar. Dorsal supraoptik komissür kesildiğinde durum oldukça farklıdır. Monoküler öğrenme sırasında sol gözden kaynaklanan sinyaller beynin yalnızca sağ yarım küresine gider. Sağ gözün monoküler testi sırasında, yalnızca daha önce görsel bilgiden yoksun olan sol hemisfere erişimi vardır ve görsel deneyim alan sağ hemisfere erişimi yoktur. Diğer bir deyişle, sağ göz eğitimsiz yarım küreye erişebilir ve eğitimli yarım küre ve burada depolanan bilgilerle hiçbir bağlantısı yoktur. Bu nedenle, dorsal supraoptik komissürde hasar olan kuşların hiçbir öğrenme transferi belirtisi göstermemesi şaşırtıcı değildir . Belki de ipsilateral yarımkürenin görsel bilgileri dorsal supraoptik komissürden başka bir yolla alabildiğine dikkat edilmelidir. Kuşlar , beynin sağ ve sol yarıkürelerini birbirine bağlayan birkaç komissural yola sahiptir (bkz. örneğin, Zeier ve Karten, 1973). Ek olarak, son zamanlarda evcil tavuklarda retinadan talamusa ve orta beyne doğru küçük bir ipsilateral projeksiyon tanımlanmıştır (O'Leary ve diğerleri, 1983). Böylece, "eğitimsiz" göz, "eğitimli" göze sunulan bilgilere bu yollardan biriyle erişebilir. Ancak, Meyer'in deneylerine bakılırsa, bu yollar boyunca aktarım gerçekleşemez.
Bu çalışmalara dayanarak tüm görsel farklılaşma becerilerinin transfer edildiğini (bkz. Zeier, 1975; Graves ve Goodale, 1979) veya dorsal supraoptik komissürün kesilmesinin bozulmamış bir durumda mümkün olan transferi her zaman bozduğunu varsaymak yanlış olur. kuş. Bununla birlikte, Meyer'in sonuçları, aktarımın belirli becerilerle gerçekleştiğini ve dorsal supraoptik komissür kesilerek önlenebileceğini kuvvetle önerir. Bu prosedürü kullanarak beynin bir yarım küresini diğerini etkilemeden "eğitmenin" mümkün olması önemlidir . Bu yöntemi, eğitimli ve eğitimsiz serebral hemisferler arasında radyoaktif urasilin RNA'ya dahil edilmesinde farklılıklar olup olmayacağını bulmak için bir damgalama durumunda kullandık (Nogi ve diğerleri, 1973).
Yumurtadan çıktıktan 1-4 saat sonra civcivlere anestezi uygulandı ve dorsal supraoptik komissür kesildi. Daha sonra civcivler 18-24 saat karanlık bir inkübatörde tutuldu. Bir gözün üzerine opak bir lastik başlık takıldı. Kısa bir süre sonra, her bir civcive radyoaktif urasil enjekte edildi ve önceki prosedüre benzer şekilde dönen, titreyen sarı ışığın önüne yerleştirildi (bkz. Şekil 2.2, K). Bununla birlikte, Şekil 1'de gösterilen maruz kalma koşulları yerine, 2.3, her civciv, eğitim için kullanılan nesneye "bakan" bir "sincap çarkına" yerleştirildi. Çarkın opak duvarları ve üzerinde tavuğun durduğu bir tel örgü zemini vardı (Şekil 3.2, L). Civciv çarkı döndürmekte serbestti, ancak ışık uyaranından yaklaşık 50 cm uzakta sabit bir konumda kaldı. Tekerlek devirleri , özel manyetik sayaçlar aracılığıyla otomatik olarak sayıldı. Civciv toplam 60 dakika cisme maruz bırakıldı.
Civcivin uyarıcı nesne için bir tercih geliştirip geliştirmediğini ve eğer öyleyse, bu tercihin beynin "eğitimli" yarımküresiyle sınırlı olup olmadığını öğrenmek gerektiğinden, damgalamanın gücünü ölçmek gerekliydi. Bu ölçüm için kullanılan aparat Şek. 3.2, B, C. Tavuk, eğitildiği çarka benzer bir çarkın içine yerleştirildi. Bu tekerlek, raylar boyunca hareket edebilen küçük bir platform üzerine monte edildi . 250 cm uzunluğundaki yolun bir ucunda “tanıdık” yanıp sönen sarı bir ışık, diğer ucunda ise dönen, yanıp sönen kırmızı bir ışık vardı (Şekil 3.2, B). Dönen tekerlek mobil platforma, tavuk nesneye yaklaşmaya çalıştığında platform ve tavuk ters yönde hareket edecek şekilde bağlandı. İletim sistemi sağlar

Resim: 3.2. Eğitim ve test için kurulumlar. A. Çıkrıktaki civciv yaklaşık 50 cm ötedeki baskı yapan bir nesneye bakıyor Çarkın opak duvarlarından biri resimde kısmen çıkarılmıştır. Tavuk tel örgü üzerinde koşar. B, Seçim testinde kullanılan aparat. C. Diyagram B'de gösterilen aparattaki çarkın parçaları. Çarkın üzerine konulan kuş bir yönde hareket etmeye çalıştığında hareketli platform ve onunla birlikte civciv ters yönde hareket eder. ( Bateson ve Wainwright'tan sonra B ve C , 1972.)
civciv 100 cm yürürken 18 cm ters yönde hareket etmiştir civciv cisme yaklaşmaya çalıştıkça cismin görünen büyüklüğü azalmış ve çekiciliğini kaybetmiştir. Bu nedenle, civcivin bir uyarana yaklaşma isteğinin başka bir uyarana olan arzusuyla dengelendiği noktayı belirlemek mümkün olduğundan , bu aparat davranışı "titrasyon" için kullanılabilir. Civciv tanıdık sarı ışığa yaklaşmaya çalıştığında diğer kırmızı ışığa doğru hareket etti. Birçok civciv bir noktada dönüp kırmızı ışığa yaklaşmaya çalıştı. Bir kuşun bir testten önce tek bir uyarana ne kadar uzun süre maruz kaldığı, bilinmeyen uyarana dönüp yaklaşmaya çalışmadan önce ondan o kadar uzaklaştığı bulunmuştur . Tüm seçim testlerinde civciv, titreşen iki ışık kaynağının ortasındaki bir tekerleğe yerleştirildi. Tekerleğin orta noktadan kat ettiği en büyük mesafe tercih indeksi olarak alınmıştır. Civciv tanıdık nesneden uzaklaşırsa gösterge pozitif olarak kabul edildi; pozitif indeks ne kadar büyükse, tanıdık nesne için tercih o kadar güçlüdür.
Otuz altı tavuk, bölünmüş yarımküre deneylerinde eğitildi, ancak biyokimyasal analiz için sadece 12 tanesi kullanıldı. Altısında sarı ışık sol gözü, diğer altısında sağ gözü etkiledi. Kuşları seçme kriterleri, nesneye görsel yönelimin varlığı ve motor bozuklukların olmamasıydı ve en önemli gösterge , ilk deneyde açık olan gözü kullanırken titreyen sarı ışık için net bir tercihin varlığıydı. testte diğer gözün izole kullanımına yönelik net bir tercihin olmaması. Bu kriterlere uymayan tavukların beyninin biyokimyasal analizi yapılmadı.
Son seçim yanıt testinden hemen sonra (ve enjeksiyondan 150 dakika sonra), kriterleri karşılayan civcivler sakrifiye edildi. Beyinleri hızla çıkarıldı, parçalara ayrıldı , donduruldu ve Bölüm 1'de açıklanan yöntemlerle analiz edildi . 2.
Eğitimin radyoaktif urasilin RNA'ya dahil edilmesi üzerindeki etkisini ortaya çıkarmak için, "eğitimli " ve "eğitimsiz" beyin yarıkürelerindeki standartlaştırılmış spesifik aktivite seviyeleri arasındaki fark hesaplandı (Şekil 3.3). Tek önemli fark, ön beyin çatısı alanında bulundu . "Eğitimli" yarım küreye dahil olma düzeyi, "eğitimsiz" yarımküreye göre ortalama %15,2 daha yüksekti. Beynin iki yarımküresinin farklı bölümlerindeki radyoaktif haberci havuzlarının boyutu önemli ölçüde farklılık göstermedi (s. 60-61). Bu gerçek, işaretli bazın makromoleküllere dahil edilmesinin, havuzun boyutundaki asimetrik değişikliklerle açıklanması olasılığını hariç tutar; örneğin, karmaşık bir görsele maruz kaldıktan sonra serebral hemisferlere kan beslemesindeki farklılıkların sonucu. uyaran (Bondy ve Morelos, 1971).
, ön beyin çatısında (Bölüm 2.1) karanlık kontrole kıyasla radyoaktif lizinin proteine daha fazla dahil edilmesi gözlemlenmiştir. bunda
120

Resim: 3.3. Eğitimli (siyah çubuklar) ve eğitimsiz (kesikli çubuklar) önbeyin çatısının yarısının, tabanının ve enine bir dorsal supraoptik komisyona sahip tavukların orta beyninin standartlaştırılmış spesifik RNA aktivitesi. Eğitimli taraf, uyarılmış gözün kontralateralidir. İçerme, sadece ön beyin çatı dokusunda değil , eğitimli tarafta önemli ölçüde daha yüksekti (P <0.02). (Horn ve diğerleri, 1971.)
Bu bölgede, etiketli urasilin RNA'ya en hızlı dahil edilmesi, görsel bir nesneye maruz bırakıldığında meydana geldi (Bölüm 2.2). Bu veriler arasındaki yakın eşleşme umut vericiydi. Bölünmüş beyinli tavuklarda yapılan çalışmalar, damgalama prosedürünün biyokimyasal sonuçlarının, öğrenme ile ilişkili spesifik olmayan etkilerle açıklanabilme olasılığını azaltmıştır . Örneğin, eğitilmiş ve karanlıkta barındırılan tavuklar arasındaki biyokimyasal farklılıklar (Bölüm 2) , lokomotor aktivite veya stres seviyesindeki farklılıklar ile açıklanabilir. Bu, bölünmüş beyinli tavuklardaki "eğitimli*" ve "eğitimsiz" yarıküreler arasındaki farkları açıklayamaz. Bununla birlikte, öğrenmenin diğer spesifik olmayan sonuçları bu kontrol deneyleri tarafından ortadan kaldırılmamıştır.
dorsal supraoptik komisyonun kesilmesi ve bir gözün kapatılmasının birleşik etkisi , beynin bir yarım küresinin görsel uyarımını tamamen durdurmak veya çok önemli ölçüde azaltmaktı . Karanlıkta tutulan tavuklarda bile görsel yollarda impuls aktivitesi kaydedilmesine rağmen, görsel uyarı ile deşarj sıklığı artar (Brown ve Nogi, 1979). Yoksun olmayan yarımkürede daha yüksek bir sinaptik aktivite seviyesi ve sonuç olarak, aracıların artan tüketimi, protein ihtiyacını artırabilir ve bu da sentezinde bir artışa yol açar. Bu nedenle, "öğrenilmiş" taraftaki ön beynin çatısında progenitör alımının artması, bilgi depolamadaki farklılıklardan ziyade hemisferlere giren impulsların sayısındaki farklılıklardan kaynaklanıyor olabilir. Benzer bir açıklama, imprinting sırasındaki biyokimyasal değişikliklerle ilgili daha önce açıklanan iki çalışma dizisinin sonuçları için de geçerlidir (Bölüm 2.1 ve 2.2). Titreşen ışığa maruz kalan civcivler ile karanlığa maruz kalan civcivler arasındaki biyokimyasal farklılıklar , uyarılmış dürtü aktivitesi seviyelerindeki farklılıklardan kaynaklanıyor olabilir ve öğrenmenin doğrudan etkileriyle hiçbir ilgisi olmayabilir.
Duyusal uyarımın bazı etkilerinin kontrolü
Yukarıda listelenen bazı güçlüklerin üstesinden gelme girişiminde, duyusal uyarımdan kaynaklanabilecek kısa vadeli değişiklikleri öğrenmenin daha kalıcı etkilerinden ayırmanın bir yolunu aradık (Bateson ve diğerleri, 1961). Bunu yaparken, çeşitli verilere göre, damgalama kuvvetinin , baskı yapan nesneye maruz kalma süresinin artmasıyla kademeli olarak arttığı gerçeğini kullandık (Sluckin ve Salzen, 1961; Connolly, 1968; Zajonc ve diğerleri, 1973; Bateson ve Jaeckel , 1976). Bununla birlikte, maruz kalma süresinin uzatılmasıyla baskı gücünün sonsuza kadar artacağı varsayılamaz. Buna göre, kümülatif öğrenme Düzeyi ile maruz kalma süresinin süresi arasındaki ilişkinin Şekil 1'de gösterilen eğri ile ifade edileceğini varsaydık . 3.4. Bu bağımlılıktan , civcivin , nesnenin ilk kez maruz kalması sırasında nesne hakkında nispeten az şey öğrendiği sonucu çıkar. Maruz kalma arttıkça, öğrenme oranı artar, böylece kümülatif öğrenme eğrisinin eğimi daha dik olur. Ardından , büyük olasılıkla nesne zaten büyük ölçüde bilindiğinden ve kümülatif öğrenme eğrisi kademeli olarak düzleştiğinden, öğrenme hızı yavaşlar. Bu eğrinin özelliği, apsis üzerindeki belirli bir "adım* A/'nın, A/'nın nerede seçildiğine bağlı olarak, ordinatta farklı "adımlara*" karşılık gelebilmesidir. Şekil düşünün. 3.4. Civcivlerin en hızlı öğrenme döneminde, kümülatif öğrenme eğrisinin eğimi en diktir. Bu dönemde, başlangıç dönemini takip eden civciv nispeten öğrenir.
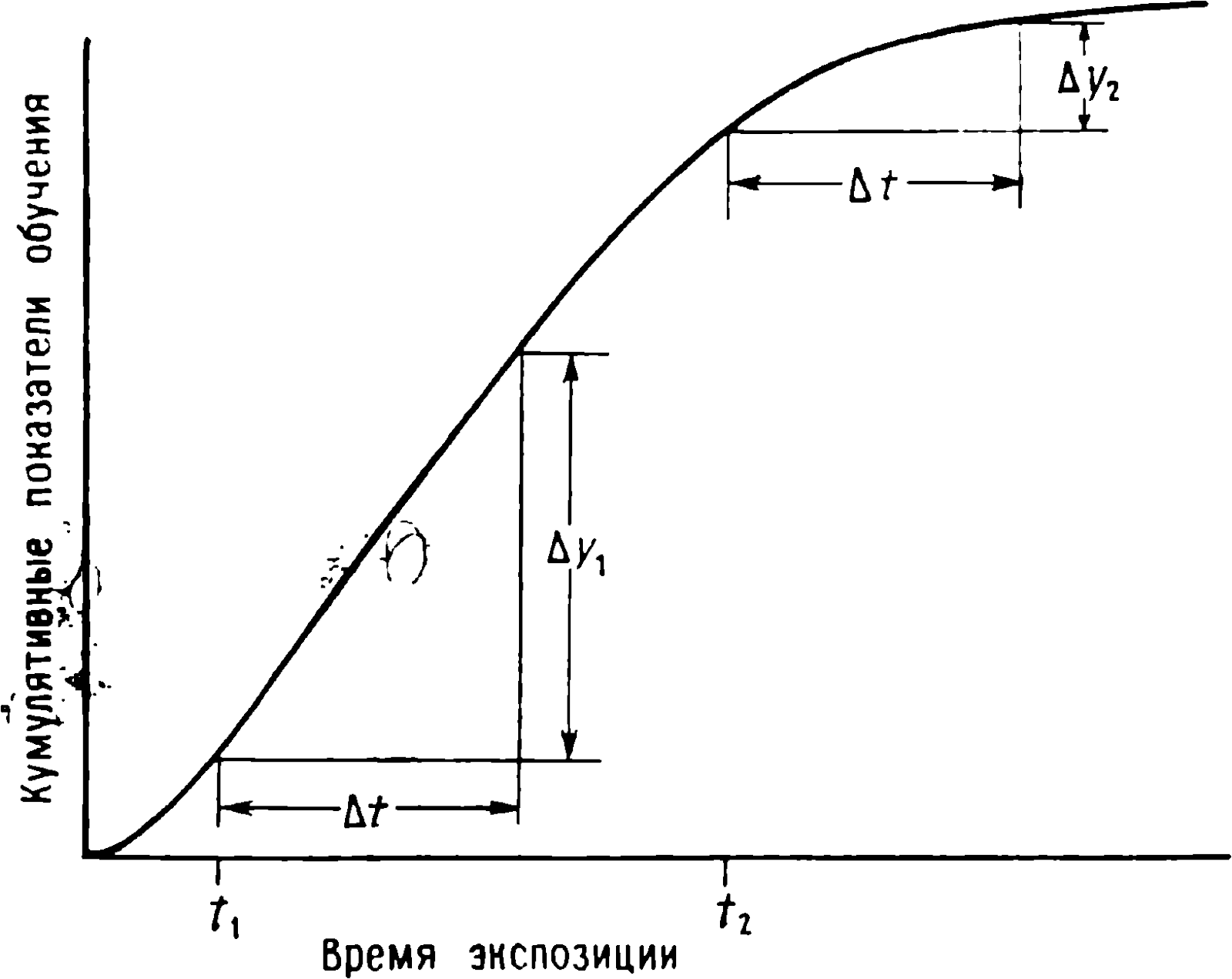
Resim: 3.4. Tahmini öğrenme eğrisi. Kümülatif öğrenme göstergelerinin görsel bir nesneye maruz kalma süresine bağımlılığının bir S şekline sahip olduğu varsayılmaktadır. Daha ayrıntılı açıklamalar için metne bakın.
D/ aralığı için büyük miktarda bilgi (D#i) . Daha sonraki / 2 periyodunda , öğrenme hızı azalmaya başladığında , civciv D/ aralığında sunulan nesne hakkında çok daha az şey öğrenir, yani D1/ 2 , D1/ i'den daha az olacaktır. Öğrenmenin nöral bağıntıları aynı eğri ile tanımlanırsa, aynı mantık geçerlidir: maruz kalma D/ periyod 6'dan sonra nöral değişiklikler t2'den daha belirgin olmalıdır .
İki grup tavuğun bir miktar uyarana maruz kaldığını varsayalım , ancak bir grup bu uyarandan t süresi boyunca etkilenirken, diğer grup daha uzun bir etkiye - t 2 süresi boyunca - maruz kaldığını varsayalım. (Şekil 3.4). Bu varsayımsal deneyde, her iki tavuk grubuna da ertesi gün etiketli urasil enjekte edilir ve aynı uyaran tekrar sunulur. Bu sefer her iki &t grubu için maruz kalma süreleri aynıdır. Tekrarlanan maruz kalma üzerine, uyaranı sadece kısa bir t' süreyle gören tavuklar ondan bilgi almaya devam etmelidir, yani D/ periyodu boyunca , bu tavuklar uyaran hakkında onu ilk kez gözlemleyen tavuklardan daha fazla şey öğrenmelidir . daha uzun bir t2 periyodunda zamanlayın ( Şekil 3.4, D#i'nin Dg/ 2'den büyük olduğu yer ). Bu nedenle, radyoaktif urasilin dahil edilmesi yalnızca öğrenme süreciyle ilişkiliyse, uyaranı bir süre gören tavuklarda dahil olma düzeyi, onu t 2 için görenlere göre daha yüksek olmalıdır \\ bu, açılma düzeyinin, damgalayıcı uyarana ilk maruz kalma süresiyle ters orantılı olması gerektiği anlamına gelir. Tersine, işaretli urasilin dahil edilmesi görsel deneyimin kısa vadeli etkisine bağlıysa , o zaman iki tavuk grubu arasında hiçbir fark olmamalıdır: radyoaktif habercinin dahil edildiği süre boyunca, her iki grup da aynı uyarıyı alır. aynı zamanda /. Bu spekülatif deney , hipotezlerin her biri için farklı tahminlerin yapılmasına izin verir . Bir yandan, eğer biyokimyasal değişiklikler spesifik olarak damgalama işlemiyle ilgiliyse, ikinci maruz kalma süresinden sonra ölçülen dahil etme A/ başlangıçta daha uzun bir süre için eğitilmiş tavuklarda daha düşük olmalıdır / 2 . Öte yandan, biyokimyasal değişiklikler görsel uyarının kısa vadeli etkisi ile ilişkilendiriliyorsa, bu durumda dahil etme her iki kuş grubunda da aynı olmalıdır.
Bu hipotezleri test etmek için tasarlanan deneylerde, iki grup yerine dört grup tavuk kullanıldı. Yumurtadan çıktıktan birkaç saat sonra, sosyal olarak izole civcivler 60 dakika boyunca yukarıdan sabit ışıkla aydınlatıldı. Bu ön uyarım, sonraki öğrenmenin etkisini arttırmak için kullanıldı (Bateson ve Wainwright, 1972). Civcivler, yumurtadan çıktıktan 24-27 saat sonra bir sincap çarkına yerleştirildi ve turuncu, yanıp sönen, dönen bir ışık kullanılarak eğitildi. Maruz kalma süresi /i , 20, 60, 120 veya 240 dakikaydı. Maruz kalma sürelerine ek olarak, civcivler karanlık inkübatörlerde izole edilmiş ve zayıf yeşil ışık altında bakılmıştır. Bu dört gruptaki yaklaşma tepkisinin şiddeti, şekil 2'de gösterilmiştir. 3.5, L. Tavuk koştuğunda çark döndü. Bu çarkın çapı 30 cm olduğu için civcivler çarkın her tam dönüşünde cisme doğru yaklaşık 94 cm koşmuşlardır. Nesne civcivler için güçlü bir çekiciliğe sahipti : 120 dakika boyunca ona maruz kalan civcivler ortalama olarak 0,5 km'den fazla koştu ve 240 dakika boyunca ona maruz kalan civcivler 1 km'den fazla koştu.
yukarıda açıklananla aynı aparat ve aynı yeni nesne kullanılarak seçim durumunda ölçülmüştür (s. 67-69). Dört grubun her biri için ortalama tercih puanları , Şekil 1 ve 2'de gösterilmektedir. 3.5B Tanıdık bir nesneyi tercih etme gücü maruz kalma süresiyle ilişkilidir . Testin ardından civcivler tekrar karanlık inkübatöre alındı.
Yumurtadan çıktıktan sonraki ikinci günde tüm civcivlerin kalp bölgesine 20 µCi radyoaktif urasil enjekte edildi ve civcivlerin tamamına geri verildi.

Resim: 3.5. Turuncu titreşen ışığa yaklaşan civcivlerin tepkilerinin ortalama değerleri ve standart hataları ve bu uyaranı tercih ettikleri seçim. A. Yaklaşma tepkisi, civcivler civcivler nesneye doğru koşarken nesneye yaklaşmadıklarında çıkrığın toplam hareket uzunluğu cinsinden ölçülür. B. Tanıdık nesne tercihi, tanıdık ve yeni nesneler arasındaki raylar boyunca orta noktadan maksimum hareket mesafesi ile ifade edilir. Pozitif bir değer, tanıdık bir nesne için bir tercihi, negatif bir değer ise yeni bir nesne için bir tercihi gösterir. Tanıdık nesne tercihi, maruz kalma süresiyle ilişkilidir (Spearman korelasyon katsayısı r ve 0,325; P<0,01). B. Yumurtadan çıktıktan sonraki ikinci gündeki ikinci maruz kalma süresi boyunca tepkiye yaklaşın. (Bateson ve diğerleri, 1973.)
yaklaşık 45 dakika boyunca karanlık bir inkübatörde. Daha sonra tekrar 60 dakika boyunca tanıdık titreşen ışığa maruz bırakıldılar ( Şekil 3.4'te Af). İkinci eğitim döneminde civcivlerin yaklaşma tepkisi, şekil 2'de gösterilmektedir. 3.5, B. Önceki gün 120 ve 240 dakika eğitilen kuşlar daha aktifti , ancak bu eğilim istatistiksel olarak anlamlı değildi. Tavuklar, enjeksiyondan 150 dakika sonra kurban edildi, Şekil 1'de gösterildiği gibi beyin diseke edildi. 2.5, L, ancak tavan ön-

beyni ayrıca ön üçte iki ve arka üçte bire bölündü. Doku parçaları donduruldu, kodlandı ve asitte çözünmeyen çökeltiye etiket katılma seviyesi belirlendi. Farklı zamanlarda analiz edilen numune serileri arasındaki değişkenliği ortadan kaldırmak için, her dahil etme indeksi , standartlaştırılmış bir spesifik aktivite olarak, yani alındığı serideki tüm numuneler için ortalama değerin yüzdesi olarak ifade edildi .
Ön beyin çatısının ön kısmı için, radyoaktif Urasil'in (Şekil 3.6, K) dahil edilmesi için önceki gün eğitim süresi ile negatif bir korelasyon bulundu. 60 ve 240 dakika çalıştırılan gruplarda ön beyin çatısının ön kısmındaki farklılıklar da istatistiksel olarak anlamlıydı (P<0.05). Beynin diğer bölümlerindeki farklılıklar istatistiksel olarak anlamlı değildi.
1. gün eğitimin 2. gün urasil'in ön beyine dahil edilmesi üzerindeki etkisi , kuşların 2. gün eğitilip eğitilmemesine bakılmaksızın ortaya çıkabilir. Örneğin, ilk eğitim döneminin bir sonucu olarak dahil etme önemli bir süre için bastırılabilir ve eğitim süresi ne kadar uzun olursa, sentez o kadar fazla bastırılabilir. Bunu test etmek için , civcivlerin kontrol gruplarının yumurtadan çıktıktan sonraki 1. günde 20, 60, 120 ve 240 dakika eğitildiği bir deney gerçekleştirildi. Ancak 2. gün radyoaktif urasil inkorporasyonu ölçüldüğünde civcivler tekrar uyarana maruz bırakılmadı, karanlıkta bırakıldı. Diğer açılardan, kontrol grupları deneysel olanlardan farklı değildi. Eğitim süresi boyunca kontrol tavuklarında yaklaşım tepkisi ve tercih tezahürü , deneysel olanlarla aynıydı. Ancak kontrol tavuklarında hiçbir grupta ve beynin herhangi bir bölümünde önemli bir değişiklik bulunmadı (Şekil 3.6, B). Bu nedenle , eğer kuşlar daha fazla eğitime tabi tutulmadıysa, 1. gündeki eğitim süresinin 2. gündeki öncü maddenin makromoleküllere dahil edilmesi üzerinde hiçbir etkisi olmamıştır. Bu sonuç, yumurtadan çıktıktan sonraki 2. günde 60 dakika eğitilen kuşlarda bulunandan keskin bir şekilde farklıdır .
eğitimin ilk 240 dakikasında tanıdık bir nesne tercihi artmaya devam ettiğinden, daha uzun süre eğitilen tavukların daha fazla bilgi aldıkları düşünülebilir (Şekil 3.5.5). Bu varsayım doğruysa, 1. gün uzun süre eğitim alan deney grubu kuşlarının 2. günde bağlandıkları nesne hakkında 1. gün kuşlarına göre daha az şey öğrendikleri de varsayılabilir. 1. gün daha kısa bir süre boyunca eğitildi.
Elbette, iyi eğitilmiş civcivlerin tanıdık bir nesneyi ikinci kez gördüklerinde, ona daha az ilgi göstermeleri ve bu nedenle 1. günde kısa bir süre için eğitilen kuşlardan daha az görsel uyarı almaları mümkündür. Ancak, 1. günde uzun süre eğitilen kuşlar, 2. günde kısa eğitim periyodundaki kuşlara göre tanıdık bir nesneye yaklaşma tepkisini daha güçlü verdiğinden, bu olasılık reddedilmelidir.
Bu deneyin sonuçları, önceki deneylerden elde edilen verilerle birlikte, özellikle "bölünmüş" beyinli tavuklar üzerinde elde edilen verilerle birlikte incelendiğinde, ön beyin çatısının ön kısmında meydana gelen biyokimyasal değişikliklerin beyin ile ilişkili olduğu görülebilir. duyusal uyarımdaki kısa vadeli değişimler gibi daha genel yan etkilerden ziyade, damgalama için gerekli öğrenme süreci .
Bununla birlikte, imprinting prosedürünün beynin ön çatısının ön kısmındaki nöronların farklılaşmasını etkilemesi mümkündür : 1. günde bu alanın gelişimi ne kadar ilerlemişse, 2. günde meydana gelen değişiklikler o kadar küçük olacaktır. deney grubu yeniden eğitilir. Böyle bir süreç kolayca hayal edilebilir. Örneğin, Valverde ( 1971) fareleri karanlıkta 20 güne kadar büyüttü. Daha sonra onları çeşitli süreler boyunca aydınlattı ve son olarak görsel korteksteki nöronların morfolojisine baktı. Bu nöronların dendritleri, dendritik dikenler adı verilen küçük çıkıntılara sahiptir. Akso-spike sinapsları oluşturan bir veya daha fazla presinaptik lifin terminallerini sonlandırırlar . Valverde , görsel korteksteki hücrelerin apikal dendritlerinin birim uzunluğu başına dendritik dikenlerin sayısını saydı. Dendritin birim uzunluğundaki diken sayısı, ışığa maruz kalma süresinin artmasıyla arttı. Prensipte aynı etki, yukarıda açıklanan deneylerde iki gün boyunca eğitilen deneysel tavuklarda da meydana gelebilir. Dikenlerin yoğunluğu, 1. günde 240 dakika maruz kaldıktan sonra beynin ön çatısındaki nöronlarda maksimuma çıkabilir . Bu durumda 2. günde müteakip maruz kalma , dikenlerin yoğunluğunda ek bir artışa yol açmayacaktır . Radyoaktif urasilin RNA'ya dahil edilmesi bu gelişimsel süreci yansıtıyorsa, o zaman 2. günde dahil etme küçük olacaktır. 1. günde örneğin 60 dakika damgalama uyaranına maruz kalan civcivler için durum farklı olacaktır. Böyle bir eğitim döneminden sonra omurga yoğunluğundaki varsayımsal artış, maksimumun altında kalabilir. Bu durumda uyaranların müteakip sunumu , dendritin birim uzunluğu başına diken yoğunluğunu arttırmalıdır ve böyle bir artış, radyoaktif urasil'in RNA'ya dahil edilmesinde karşılık gelen bir artışla ilişkilendirilmelidir. Bu nedenle, öncünün bu gruptaki tavuklara dahil edilmesi, 1. günde *40 dakika süreyle uyarılanlardan daha fazla olmalıdır. Sonraki deneyler (Bradley ve diğerleri, 1981; Nogi ve diğerleri, 1985a, b), dendritik dikenlerin yoğunluğu üzerinde Rint uyarısının böyle bir etkisini doğrulamasa da , genel olarak bu argüman önemli kabul edilmelidir. Bu nedenle, yukarıdaki çalışmanın sonuçlarının yorumlanmasını açıklığa kavuşturmak için aşağıdaki deneyler (Bateson ve diğerleri, 1975) yapılmıştır .
Biyokimyasal ve Davranışsal Parametrelerdeki Bireysel Varyasyonlar
Bir grup tavuğa belirli bir süre için bir damgalama nesnesi sunulduğunda, tanıdık bir nesne için ortaya çıkan tercihin geniş bir şiddet aralığı gözlenir. Ek olarak, beyne dahil edilen radyoaktif urasil miktarında da eşit derecede geniş bir varyasyon vardır. Davranışsal ve biyokimyasal parametrelerin bu doğal dağılımı bizim tarafımızdan özel olarak incelenmiştir. Asitte çözünmeyen fraksiyonlara etiketli urasil eklenmesi baskı işlemiyle yakından ilişkiliyse, o zaman daha güçlü bir tercih geliştiren civcivlerde daha yüksek öncül birleştirme düzeyi olmalıdır. Açma, uzun süreli stimülasyon nedeniyle nöronların spesifik olmayan gelişimi ile ilişkiliyse , o zaman tüm tavukların aktif durumda olduğu göz önüne alındığında, hepsi aynı anda uyarana maruz kaldığı için aralarında herhangi bir yön farkı olmamalıdır. .
Bu deneylerde 106 civciv kuluçkadan çıktıktan sonraki 1. günde etiketli urasil uygulandı. Daha sonra 72 dakika boyunca sarı (Şekil 2.2, L) veya kırmızı (Şekil 2.2, B) dönen titreyen ışıkla sunuldu. Yukarıda açıklanan deneylerde olduğu gibi , tavuklar, baskı yapan bir uyaranın önünde dönen bir çarkta eğitildiler. Eğitimden 20 dakika sonra, tanıdık ve yeni titreyen ışık arasındaki seçim tepkisi özel bir aparat içinde test edildi (Şekil 3.2, B). Her bir civcivin davranışının çeşitli göstergeleri ölçüldü ve bunlar daha sonra tablo haline getirildi (Tablo 3.1). Ölçümden sonra tavuklar kesildi, beyin yukarıda tarif edildiği gibi parçalara ayrıldı (Şekil 2.5, L), ek olarak ön beyin ön ve arka kısımlara ayrıldı ve radyoaktif urasil bu beyinden asitte çözünmeyen fraksiyonlara dahil edildi bölgeler ölçülmüştür.
tek doğrudan ölçüsü olan tanıdık uyaran tercihi, progenitörlerin rostral ön beyin çatısına dahil edilmesiyle pozitif olarak ilişkiliydi; kuşlar ne kadar iyi eğitilirse, yalnızca bu bölgede urasilin RNA'ya dahil edilmesi o kadar yüksek oluyordu. Ancak bu korelasyon zayıftı (Tablo 3.2). Tanıdık bir uyaran tercihi ile radyoaktif maddenin dahil edilmesi arasındaki bağımlılığın olması mümkündür.
Tablo 3.1. Bateson, Horn ve Rose'un deneylerinde kaydedilen davranışsal göstergelerin listesi (Bateson ve diğerleri, 1975)
Tablo 3.2. Dört beyin bölgesindeki radyoaktif urasilin asitte çözünmeyen maddelere dahil edilmesi ve davranışı arasındaki korelasyonlar; Spearman korelasyon katsayıları verilmiştir (Bateson ve diğerleri, 1975)
* P<0.05; ** P<0.001; P<0.05.
Öğrenme yaklaşımı gecikmesi ile urasil birleşimi arasındaki pozitif korelasyon, civciv ne kadar yavaş yaklaşmaya başlarsa, RNA'ya katılımın o kadar yüksek olduğunu gösterir. İlk yaklaşma tepkisi ile negatif bir korelasyon, civciv ilk 20 dakika içinde ne kadar çok yaklaşırsa, orta beyin hücrelerinin RNA'ya dahil edilmesinin o kadar düşük olduğunu gösterir.
Beynin ön kısmındaki yüksek urasil başka bir faktör tarafından kısmen maskelenebilir. Öğrenme sırasında uyarana hızlı bir şekilde tepki veren kuşlarda , beynin tüm kısımlarında habercinin düşük düzeyde dahil olduğu gözlemlendi (Tablo 3.2). İyi telaffuz edilmiş bir yaklaşma tepkisine sahip kuşlar, eğitilebilir en iyi kuşlar olarak kabul edilebildiğinden (Bateson ve Jaeckel, 1974), ön beyinde yer alma iki karşıt etkiden kaynaklanabilir. Bunlardan biri beynin tüm bölgelerinde ortak olan asitte çözünen fraksiyonda inklüzyonda azalmaya yol açabilirken, diğeri anterior çatıya özgü bu alanda inklüzyonda artışta ifade edilebilir. Eğitim sırasındaki yüksek aktivite seviyelerinin beklenen genel olumsuz etkisi, önbeyin çatısının iki bölgesi için spesifik radyoaktivitenin, önbeyin tabanının yanı sıra orta beyindeki spesifik radyoaktivitenin bir yüzdesi olarak ifade edilmesiyle ortadan kaldırıldı. kuş. İki ön beyin çatı bölgesi için katılım puanları ile sekiz davranışsal puan arasındaki korelasyonlar Tablo 1'de gösterilmektedir. 3.3. Tek
Tablo 3.3. Davranış özellikleri arasındaki ilişkiler
ve 116 tavukta ön beyin çatısının ön ve arka bölümlerine radyoaktif urasil eklenmesi; Spearman korelasyon katsayıları verilmiştir (Bateson ve diğerleri, 1975)
* PCO.OOl.
önbeyin çatısının rostral kısmına bir etiket eklenmesi arasındaki ilişki için istatistiksel olarak anlamlı bir korelasyon katsayısı bulundu . Bu korelasyon pozitiftir ve oldukça anlamlıdır (Şekil 3.7). Başka bir deyişle, tüm davranışsal göstergeler içinde, yalnızca damgalamanın tezahürü, radyoaktif urasilin rostral ön beyindeki RNA'ya dahil edilmesiyle pozitif olarak ilişkilidir. Tanıdık bir uyaran tercihi ile öncünün arka ön beyine dahil edilmesi arasındaki ilişki istatistiksel olarak anlamlı değildi.
Bu deneylerin sonuçları, biyokimyasal değişikliklerin, öğrenmenin nöronların büyümesi ve gelişmesi üzerindeki bazı genel etkilerinin sonucu olduğu hipotezini destekliyor mu? Bu hipotezden, radyoaktif urasilin dahil edilmesindeki sistematik farklılıkların, uyarana maruz kaldıkları süre boyunca aktif olmaları durumunda farklı kuşlarda ortaya çıkmayacağı sonucu çıktı. Bu durum önemlidir. Eğer bazı civcivler

Standartlaştırılmış Öncül İçerme İlişkileri
Resim: 3.7. Tanıdık nesne tercihinin göstergeleri ile radyoaktif urasilin rostral ön beyin çatısından asitte çözünmeyen fraksiyonlara dahil edilmesi arasındaki korelasyon. Farklı civciv grupları ve farklı eğitim koşulları (sarı veya kırmızı baskı) arasındaki farkları ortadan kaldırmak için tercih puanları standardize edilmiştir . Belirli bir kuşun ön beyninin çatısının spesifik aktivite değerleri, aynı kuş için orta beynin ve ön beynin tabanının ortalama spesifik aktivitesine bölündü. Bu standartlaştırılmış yüzdeler daha sonra sıralandı ve 22 civciv içeren 5. grup hariç, grup başına 21 civciv içeren beş gruba ayrıldı . Spearman'a göre ortaya çıkan korelasyon katsayısı, 0.316'ya eşit, P<0.001'de yüksek bir güvenilirliğe sahipti. (Bateson ve diğerleri, 1975.)
Uykulu olsalardı, gözleri kısmen veya tamamen kapalı olsaydı, uyanık civcivlere göre daha az retinal uyarı alırlardı ve sonuç olarak görme yollarında daha az aktivite olurdu. Ek olarak, fizyolojik süreçler, davranıştaki farklı aktivasyon ve uyanıklık seviyelerinde farklı şekilde ilerler . Bu seviyelerdeki dalgalanmaların , en azından bazı memelilerde, görsel yollardaki uyarılmış aktivite iletimindeki değişikliklerle ilişkili olduğuna dair kapsamlı kanıtlar vardır (bkz. Hoot, 1976; Wurtz ve diğerleri, 1980; Rizzolatti, 1983); kuşlarda da benzer mekanizmaların çalıştığı varsayılmaktadır. Görsel deneyimin çeşitli koşullarının görsel korteksteki nöronların yapısını etkilediği bilinmektedir (s. 77; Lund tarafından yapılan incelemeye bakın, 1978). Sonuç olarak, civciv ne kadar aktif olursa, görsel dürtülerin ön beyne akışının o kadar yüksek olacağı tartışılabilir. Bu yüksek seviyedeki ani yükselme aktivitesi, nöron büyümesini ve RNA sentezini güçlü bir şekilde uyarabilir . Aktif civcivlerin daha yüksek tercih yanıtı oranlarına sahip olması beklenebileceğinden, tercih ile emsalin dahil edilmesi arasında bir korelasyon olmalıdır.
140 çırpma. Bununla birlikte, aktivasyon seviyesi, etiketli urasil'in RNA'ya dahil edilmesini etkiliyorsa, habercinin dahil edilmesi ile diğer bazı davranışsal özellikler arasında da bir korelasyon olmalıdır. Örneğin, tanıdık bir uyarana yaklaşmanın tepki gecikmesi, civcivin bir nesneye ne kadar hızlı tepki verdiğini yansıttığı için aktivasyon seviyesinin bir göstergesidir. Bununla birlikte, eğitim ve test durumundaki gizli dönem ile öncünün ön beyine dahil edilmesi arasında bir ilişki yoktur. Aslında, öğrenme dönemindeki davranış özelliklerinin hiçbiri kaynaştırma süreciyle ilişkili değildir (Tablo 3.3). Bununla ve diğer tepkisellik göstergeleri, uyarılma veya kaygı düzeyi arasında bir korelasyon yoktu. Bu sonuçlar, tavuk davranışının listelenen yönlerinden hiçbirinin ön beyinde meydana gelen değişikliklerin nedeni olamayacağını göstermektedir. Bu nedenle, genellikle damgalama sırasında bilgi izlerinin kaydedilmesine eşlik eden çoklu faktörleri ayırmak için doğal davranışsal değişkenlik kullanılmıştır.
Yorum
biyokimyasal çalışmasının üç ana amacı vardır: 1) civciv damgalandıktan sonra beyinde biyokimyasal değişikliklerin meydana gelip gelmediğini belirlemek. Bu tür kaymalar bulunursa, o zaman 2) önemsiz bir açıklamaları olup olmadığını öğrenin ve 3) bu biyokimyasal değişiklikleri en azından oldukça kaba bir biçimde yerelleştirmeye çalışın. İlk hedefe ulaşıldı (bölüm 2). İkinci soruyla ilgili olarak , rostral ön beyin çatısında hızla meydana gelen biyokimyasal değişikliklerin öğrenmenin çeşitli yan etkilerine bağlı olamayacağını gösteren veriler yavaş yavaş elde edildi. Üçüncü soru ile ilgili olarak, ilk çalışmalar , değişikliklerin ön beyin çatısında meydana geldiğini gösterdi ve sonraki çalışmalar , bu yapının rostral üçte ikisinde daha dar bir bölge ortaya çıkardı . Böylece, üçüncü sorunu çözmedeki başarı mütevazıydı. Ancak biyokimyasal değişimler beynin belli yapılarıyla sınırlıysa, o zaman bu yapıların ne olduğunu ortaya koymak gerekir. Daha önceki tartışmalarımızdan da görülebileceği gibi (s. 48-49), çalışmanın başarısı ancak böyle bir yerelleştirme varsa mümkündür.
Özet
Bölüm'de açıklanan biyokimyasal değişiklikler. 2 eğitim prosedürünün çeşitli yan etkilerinin sonucu olabilir . Damgalayıcı bir uyaranla sunulan civcivler , kontrol grubuna göre daha ajite, daha aktif ve daha fazla duyusal uyarı alabilirler. Bu bölüm, listelenen çeşitli faktörlerin rolünü test etmek için tasarlanmış üç deneyi açıklamaktadır. 1. Dorsal supraoptik komisyonun kesilmesiyle, beynin bir yarımküresini "eğitmek" mümkün oldu. İşaretli urasil'in önbeynin çatı bölgesindeki RNA'ya dahil edilmesinin "eğitimli" yarımkürede "eğitimsiz" yarımküreden daha yüksek olduğu ortaya çıktı . 2. Görsel stimülasyonun kısa vadeli etkisinin önemi, farklı civciv gruplarının yaşamın 1. gününde farklı uzunluklarda ve ertesi gün eşit sürelerde eğitildiği bir deneyde test edildi. Sonuçlar , duyusal stimülasyonun kısa vadeli etkilerinin gözlemlenen biyokimyasal değişiklikleri açıklayabileceği önerisini desteklemedi . 3. Ön beyin çatısının rostral üçte ikisine dahil edilen radyoaktif urasil miktarı, damgalama gücünün bir göstergesi olan tanıdık bir nesneyi tercih etmenin ciddiyeti ile ilişkiliydi, ancak diğer davranışsal göstergelerle ilişkili değildi. Tavuk. Biyokimyasal değişikliklerin bilgi kaydetme ve depolamanın bazı yönlerini yansıtma olasılığı, bu kontrol deneyleri tarafından çürütülmedi, ancak diğer bazı açıklamalar onlar tarafından ortadan kaldırıldı.
EDEBİYAT
Bateson R. R. G., Jaeckel J. B. Damgalama: eğitim ve test sırasında civcivlerin aktiviteleri arasındaki korelasyonlar, Anim. Behav., 1974, 22, 899-906.
Bateson PPG t Jaeckel J. B, Chicks*'in farklı maruz kalma sürelerinden sonra tanıdık ve yeni göze çarpan nesneler için tercihleri, Anim. Behav., 1976, 24, 386-390.
Bateson PPG, Wainwright AAP Daha önce ışığa maruz kalmanın evcil civcivlerde baskı süreci üzerindeki etkileri, Behavior, 1972, 42, 279-290.
Bateson PPG, Rose SPR, Horn G. damgalama: civciv beynine urasil entegrasyonu üzerinde kalıcı etkiler. Bilim, 1973, 181, 576-578.
Bateson P, PG, Horn G., Rose SPR imprinting: [HC] urasil'in civciv beynine dahil edilmesi ve davranışı arasındaki korelasyonlar , Brain Res., 1975, 84, 207-220.
Bondy S. C., Morelos BS Uyaran yoksunluğu ve serebral kan akışı, Exp. Neurol., 1971, 31, 200-206.
Bradley P., Horn G., Bateson P. damgalama: civciv hiperstriatum ventrale'nin elektron mikroskobik çalışması, Exp. Brain Res., 1981, 41, 115-120.
Brown MW, Horn G. Civciv beynindeki nöronal plastisite: görsel deneyimin hiperstriatal nöronlar üzerindeki elektrofizyolojik etkileri, Brain Res., 1979, 162, 142-147.
Bryan R, N. Bliss EL, Beck EC üridin birleşimi. Stres sırasında fare beyin RNA'sına 8 H, Fedn. Proc. Fedn. Am. soslar. tecrübe. Blok., 1967, 26, 709.
Connolly K. evcil civcivlerde eğitim miktarının bir fonksiyonu olarak imprinting ve aşağıdaki yanıt, Quart. J. exp. Psychol., 1968, 9, 453-460.
6" Cowan WM, Adatnson L., Powell TP S. Kuş görsel sisteminin deneysel bir çalışması, J. Anat., 1961, 95, 545-563.
Cuenod M., Zeier H. Görsel interhemisferik ve komissurotomie chez le güvercini aktarın. Archs, Neurol, Neurochirg'i suisse eder. Psikiyatri." 1967, 100, 365-380.
Graves JA, Goodale MA Güvercinlerde eğitim koşulları interoküler transferi etkiler mi? İçinde: Serebral komisyonların yapısı ve işlevi (eds. I. Steele Russell, MW van Hof ve G. Berlucchi), 1979, s. 73-86, Macmillan , Londra.
Boynuz G. Dikkat ve uyarılmanın fizyolojik çalışmaları. İçinde: Bilinçli davranış için sinyallerin iletim mekanizmaları (ed. T. Desiraju), 1976, s. 285-299, Elsevier, Amsterdam.
Horn G., Horn ALD, Bateson PPG, Rose SPR "Bölünmüş beyinli" civcivde urasilin beyin RNA'sına dahil edilmesi üzerindeki baskılamanın etkileri, Nature, Londra., 1971, 229, 131-132.
Horn G., Rose SPG, Bateson PPG Bozulmamış ve "bölünmüş beyinli" civcivlerin beyinlerine tritiye urasilin monoküler damgalama ve bölgesel dahil edilmesi, Brain Res., 1973, 56, 227-237.
Horn G., Bradley P., McCabe BJ Damgalamanın morfolojik bağıntıları, Adv. davranış. Biol., 1985a, 28, 21-27.
Horn G., Bradley P., McCabe BJ Öğrenme ile ilişkili sinapsların yapısındaki değişiklikler, J. Neurosci., 1985b, 5, 3161-3168.
Hyden H. Büyüme ve fonksiyon sırasında sinir hücrelerinde protein metabolizması, Açta fizyol. tarama, 1943, 6, Ek. 17:1-136.
Jakoubek B., Horâckovâ M., Gutman E. Spinal motor nöronlarda ve siyatik sinirde artan fonksiyonel aktivite sırasında protein metabolizmasındaki değişiklikler, Proc. Int. Union Physiol. Sci., 1968, 7, 2150.
Jakoubek B., Semiginovsky B., Krauss M., Erdossovâ R. Beklenti stresi ve ACTH tarafından indüklenen beyin korteksinin protein metabolizmasının değişmesi, Life Sci., 1970, 9, 1169-1180.
Jakoubek B., Buresovâ M., Hâjek Etrychovâ J., Pavlik A., Dedicova A. ACTH'nin farelerin sinir sisteminde hızlı etiketlenmiş RNA'nın sentezi üzerindeki etkisi, Brain Res., 1972, 43, 417-428.
Lund R. D. Beynin gelişimi ve plastisitesi, Oxford University Press, New York, 1978.
Meier RE Interhemispharischer Transfer vizörü Zweifachwahlen bel komissurotomierten Tauben, Psychol. Forsch., 1971, 34, 220-245.
Myers RE İnteroküler transferde korpus kallozumun işlevi, Brain, 1956, 79, 358-363.
Myers RE, Henson CO Şempanzede taktuokinestetik öğrenmenin transferinde korpus kallozumun rolü, Archs Neurol. Psychiat., Chicago, 1960, 3, 404-409.
Myers RE, Sperry RW Optik kiazma ve korpus kallosumun kesitinden sonra kedilerde görsel form ayrımcılığının interoküler aktarımı, Anat. Kayıt." 1953, 115, 351-352.
O'Leary DDM, Gerfen CR, Cowan WM Civcivde ipsilateral retinofugal projeksiyonun gelişimi ve kısıtlanması, Dev Brain Res., 1983, 10, 93-109.
Perisic M., Mihailovic J., Cuenod M. Güvercinlerde ( Columba livla) t Int . J. Neurosci., 1971, 2, 7-14.
Rizzolatti G. Memelilerde seçici dikkat mekanizmaları. İçinde: Omurgalı nöroetolojisindeki gelişmeler (eds. J.-P. Ewert, RR Capranlca ve DJ Ingle), 1983, s. 261-295, Plenum Press, New York.
Sluckin W., Salzen EA damgalama ve algısal öğrenme, Quart. J. exp. Psychol., 1961, 13, 65-77.
Valverde F. Fare Beyin Res. 1971, 33, 1-11.
Wagle SR Amino asitlerin proteine dahil edilmesi üzerinde büyüme hormonu, kortizol ve insülinin etkisi, Archs Biochem., 1963, 102, 373-378.
Wurtz RH, Goldberg ME, Robinson DL Maymundaki görsel tepkilerin davranışsal modülasyonu: dikkat ve hareket için uyaran seçimi, Prog. Psychobiol. fizik. Psychol., 1980, 9, 44, 83.
Zajonc RB, Reimer D. L, Hausser D. Sadece tekrarlanan maruz kalma ile civcivlerde baskı ve nesne tercihinin gelişimi, J. comp. fizik.
Psychol., 1973, 83, 434-440.
Zeler H. Interhemisferik etkileşimler. İçinde: Kuşlarda davranışın sinirsel ve endokrin yönleri (Eds. P. Wright, PG Caryl ve DM Vowles), 1975, s. 163-180, Elsevier, Amsterdam.
Zeler H., Karten HJ Güvercinlerde (Columba livia), ön komissürün bağlantıları , J. comp. Neurol., 1973, 150, 201-216.
4. BÖLÜM
BEYİNDEKİ LOKALİZASYON
Tanınmış nöropsikolog Carl Lashley, kendisinin de belirttiği gibi, yıllarını "engramı aramaya" adadı. Hayatının sonlarına doğru şöyle yazmıştı: "Bazen, bellek izlerinin yerelleştirilmesiyle ilgili verilerin toplamına baktığımda, öğrenmenin kesinlikle imkansız olduğunu hissediyorum" (Lashley, 1950, s. 477-478). Bu alaycı ifadenin dayandığı verilerin bir kısmı, Lashley'nin sıçan labirenti öğrenimi üzerine çalışmasından alınmıştır. Öğrenilmiş tepkilerin uygulanmasıyla ilgili olarak, serebral korteksin farklı bölümlerinin birbirine eşdeğer olduğuna inanıyordu (Lashley, 1929). Önemli olan, çıkarılan bölgenin lokalizasyonu değil, korunmuş kabuğun hacmiydi.
Bu ideolojik mirasa rağmen, 1960'larda bağımlılıkla ilgili nörofizyolojik çalışmalarda, tepkilerin zayıflamasının, tekrarlanan uyarıyı alan duyu alıcıları ile tepkinin uygulanmasında yer alan kaslar arasında yer alan tüm sinaptik temaslarda meydana gelmediği gösterilmiştir. Bu yol boyunca yalnızca belirli bağlantılarda iletim kesintiye uğradı . Başka bir deyişle, en azından bu basit öğrenme biçiminde, nöronal değişikliklerin yerel olduğuna, yani sinir dokusunun bir bölümünün diğerine eşdeğer olmadığına dair kanıtlar birikiyordu. Aynı sonuç , baskı üzerine yapılan çalışmalardan da çıkıyor gibi görünüyor. Bu süreçle yakından ilişkili biyokimyasal değişikliklerin , incelenen beynin çeşitli bölgelerinde eşit olmayan bir şekilde lokalize olduğu ortaya çıktı. Ön beyin çatısında , daha doğrusu bu bölgenin rostral üçte ikisinde sürekli değişiklikler tespit edildi.
Ön beynin rostral kısmı oldukça geniş bir alandır. Milyonlarca nöron içerir ve anatomik olarak farklı birkaç bölüm içerir. Radyoaktif urasilin dahil edilmesindeki kaymalar , ön beyin çatısının tüm rostral kısmına eşit olarak dağılmış mı, yoksa onu oluşturan alanlardan herhangi birinde mi lokalize edilmiş ? Damgalama çalışmasında (Bölüm 2) kullanılan yöntemler, basit bir nedenden dolayı bu soruyu yanıtlamak için çok az işe yaradı: bu, rostral ön beyin çatısını küçük parçalara ayırmayı, her birini kavramsallaştırmayı ve öncül etiketli içermeyi ayrı ayrı ölçmeyi gerektirecekti. Bu karmaşık prosedür , incelenen her civciv için yapılmalıdır. Bu sorunu çözmek için farklı bir yaklaşım gerekiyordu. Beynin histolojik yapısını korurken aynı zamanda radyoizotopu tespit etmeyi mümkün kıldığı için otoradyografi yönteminin uygun olduğu ortaya çıktı .
Radyootografik yöntemi kullanmak için dokuya bir radyoizotop enjekte edilir. Daha sonra yapıyı korumak ve ince dilimler yapmak için düzeltmeniz gerekir (ayrıca önce dilimleri yapıp sonra düzeltebilirsiniz). Kesitler karanlıkta fotoğraf emülsiyonu ile kaplanır. Bir radyoizotop bozunduğunda yüklü parçacıklar yayar. Aşağıda açıklanan deneylerde, radyoaktif karbon 14C ile etiketlenmiş bileşikler kullanıldı.Bu izotop, trityum gibi, negatif yüklü beta parçacıkları yayar. Cam slaytlara yerleştirilen ve fotoğrafik emülsiyonla kaplanan doku kesitleri birkaç gün hatta aylarca karanlıkta bırakılır. Dokularda emülsiyonla temas halinde olan izotoplar , bazıları emülsiyona giren ve gümüş kristallerini bombardıman eden beta parçacıkları yayar. Daha sonra geliştirildiğinde, bu kristaller görünür gümüş taneciklere dönüşür.
radyoizotopları ışık ve elektron mikroskobu yoluyla lokalize etmek için kullanılabilir . Bununla birlikte, bu hücresel ve hücre altı lokalizasyon , rostral ön beyindeki hücre sayısı çok yüksek olduğundan, baskı çalışmaları için çok yüksek bir çözünürlük sağlar. Bu beyin bölgesinin herhangi bir bölümündeki izotop inklüzyonunun lokalizasyonunu saptamak için kullanılabilecek bir "makroradyootografik" yönteme ihtiyacımız vardı . Bu şekilde değişikliklerin gerçekten lokalize olduğu tespit edilirse, hücresel lokalizasyonlarını belirlemek için daha ince radyootografik yöntemler uygulamak mümkün olacaktır. Makroradyootografi, bazı metodolojik farklılıklar olmasına rağmen, mikroradyootografi ile aynı prensiplere dayanmaktadır . Örneğin, makroradyografide, slaytlar üzerindeki doku kesitleri genellikle bir emülsiyona daldırılmak yerine bir parça x-ışını filmi ile kaplanır. X ışını filmi üzerinde bir emülsiyon bu amaç için mükemmeldir. Işık ve elektron mikroskobu için kullanılan nükleer emülsiyonlarla karşılaştırıldığında , X ışını emülsiyonları nispeten büyük parçacık boyutlarına sahiptir. Sonuç olarak, filmin çıplak gözle görülebilen koyulaşmasına neden olmak için birim alan başına daha az sayıda gümüş tanesi yeterlidir.
Resim: 4.1. Radyo imzası için materyal hazırlamanın bazı aşamalarının şeması. Kalp bölgesine radyoaktif urasil enjekte edildi ve ardından tavuğa damgalayıcı bir uyaran verildi. Enjeksiyondan 2.5 saat sonra tavuk kesildi , beyni çıkarıldı ve donduruldu. 15 μm (L>) kalınlığında kesitler alınarak cam lamlara (B) yerleştirildi ve üzerine x-ray filmi yerleştirilmiş kesitler karanlıkta belli bir süre bekletildi ve ardından film geliştirildi.
Radyoaktif urasil kullanarak araştırma
Bir tavuğun beynine uygulanan tarif edilen yöntem, Şekil 1'de gösterilmektedir. 4.1. 14 C-urasil kalp bölgesine enjekte edildi. Eğitimden sonra, beyin daha önce olduğu gibi parçalara ayrılmadı (Şekil 2.5, L), ancak donduruldu ve kesildi. Cam lamlar üzerine yerleştirildiler, karanlıkta X-ışını filmiyle kaplandılar ve birkaç hafta karanlıkta bırakıldılar. Film daha sonra geliştirildi. Filmin koyulaşma derecesi, radyoaktivitenin bir göstergesi olarak görev yaptı: filmin alanı ne kadar koyu olursa, beyin diliminin karşılık gelen kısmındaki radyoaktivite o kadar yüksek olur (Şekil 4.4). Filmin belirli bir bölümünün koyulaşma derecesi, filmin küçük bir ışık noktası üzerine yerleştirilmesi ve bu alanın optik yoğunluğunun ölçülmesiyle tahmin edildi. Optik yoğunluk , film üzerindeki bir görüntüden geçerken ışığın yoğunluğunun ne kadar azaldığını gösterir.
Bir film üzerindeki bir görüntünün optik yoğunluğu, yüksek bir doğruluk derecesi ile belirlenebilir. Bu nedenle, ilke olarak, damgalama ve kontrol grubuna tabi tutulan tavukların beyninin belirli bir bölgesine radyoaktif habercinin dahil edilmesindeki küçük farklılıklar bile tespit edilebilir. Dezavantajı , farklılıkların oldukça basit nedenlerden dolayı belirgin olabilmesidir. Otoradyografi için doku hazırlanırken histolojik yapı korunacak şekilde sabitlenir. Fiksasyon sürecinde, makromoleküllerin çökelmesi meydana gelir ve bunun sonucunda dokuda belirli bir pozisyonda stabilize olurlar . Fiksasyondan sonra, doku artan konsantrasyondaki alkollere daldırılarak dehidre edilir. Genellikle bu prosedürün dokudan küçük molekülleri uzaklaştırdığına ve geriye makromoleküllerin kaldığına inanılır. Deneycinin ilgilendiği tek şey, etiketli habercinin niteliksel dağılımı ise, bu uzlaştırılabilir. Ancak bu, etiketin dağılımının nicel değerlendirmesinde önemlidir. Bazı fiksatifler, çözünür öncüleri sabit ve çökeltilmiş makromoleküllere bağlama yeteneğine sahiptir . Fiksatifin bu tür bir etkisinin sonucu, hatalı bir şekilde makromoleküllere dahil edilmiş gibi görünecek olan öncül miktarında bir artış olacaktır . Diğer bir zorluk ise bazı fiksatiflerin RNA ve protein kaybına neden olmasıdır (Rogers, 1973). Bu kayıpların sonucu, dokudaki işaretli makromoleküllerin sayısında bir azalma olacaktır. Bu fiksasyon artefaktlarının her ikisi de - radyoaktif öncülerin dahil edilmesindeki belirgin artış veya azalma - kantitatif otoradyografide önemli hatalara neden olur.
Bu metodolojik kusurları ortadan kaldırmak için, radyoaktif urasil'in beynin asitte çözünen maddelerine dahil edilmesini güvenilir bir şekilde yansıtan makroradyootografların hazırlanması için koşullar geliştirildi (Nogi ve McCabe, 1978). Modifikasyonumuzda, radyoaktif urasil enjeksiyonundan 2.5 saat sonra tavuk beyni çıkarıldı. Daha sonra beyin donduruldu ve -30°C'de kesitler hazırlandı. Kesitler lamlara yerleştirildi, trikloroasetik asit içine daldırıldı ve ardından distile su ile yıkandı. Makromolekülleri çökeltmek (Bölüm 2) ve yerlerine sabitlemek için asit daldırması gerçekleştirildi. Radyoaktif urasil içeren iyonlar ve küçük moleküller su ile yıkanarak kesitlerden uzaklaştırıldı. Yöntem , bu hedeflerin her ikisinin de başarılı bir şekilde gerçekleştirilmesini sağlamıştır . Bu şekilde işlenen bölümler , asitte çözünen maddelerde radyoaktiviteyi tutmadı, ancak aynı zamanda asitte çözünmeyen maddelerden radyoaktivite kaybı olmadı.
Bu metodolojik zorlukları çözdükten sonra, damgalama sırasında meydana gelen biyokimyasal değişikliklerin olası serebral lokalizasyonunu incelemeye devam edebiliriz (Nogi ve diğerleri, 1979). Radyootografik yöntem çok zaman gerektirdiğinden, yetersiz eğitimli tavukların beyinlerinin araştırma için alınması istenmeyen bir durumdu. Aktif civcivler genellikle aktif olmayan civcivlerden daha hızlı öğrenirler. Bu avantajdan yararlanmak için, aktif civcivleri seçmek için bir Yöntem geliştirmek gerekliydi. Gruplar arasındaki radyootografik farklılıkları yorumlarken herhangi bir şüpheye mahal vermemek için hem deney hem de kontrol grupları için tavukların bu yöntemle seçilmesi gerekiyordu . Örneğin , aktif davranışa sahip damgalanmış civcivlerin beyinleri , karanlıkta tutulan rastgele seçilmiş (dolayısıyla dahil edilmiş ve aktif olmayan) civcivlerin beyinleriyle karşılaştırıldığında, iki grubun beyin durumundaki farklılıklar daha yüksek aktiviteden kaynaklanıyor olabilir. damgalanmış civcivlerin. , ve bu şekilde damgalamanın kuvvetleri değil. Bu varsayım sadece teorik değildir: urasilin orta beyindeki makromoleküllere dahil edilmesi, yaklaşım faaliyetinin seviyesine bağlıdır (Tablo 3.2).
ve ertesi gün eşit süreler için eğitildiği bir deneye dayanıyordu (Bölüm 3.2). Aynı zamanda, tavuklar seçilebilir ve 1. gündeki aktivitelerine göre benzer çiftler halinde gruplandırılabilir. Deneyin koşulları , 1. günde en uzun süre eğitilen civcivlerin 2. günde en düşük selef katılımına sahip olması gerektiğini tahmin etmeyi de mümkün kıldı (Şekil 3.6, L).
Başlangıçta, tüm civcivler eğitimden önce 30 dakika boyunca yukarıdan beyaz, sabit ışığa maruz bırakıldı (s. 73). Daha sonra her civciv, bir damgalama uyaranının önüne dönen bir tekerleğe yerleştirildi . Sarı ışıkla aydınlatılan yatay bir yuvaya sahip hareketli bir silindirdi . Yarık pencerede görülüyordu ve saniyede dört kez yukarı doğru dönüyordu. Bu titreyen ışığın üzerinde, bir ev tavuğunun annelik çağrısının verildiği bir hoparlör vardı. Bu ek uyaran, hareket eden bir nesneyi takip etme eğiliminin, nesne bu türün annenin çağrı özelliğini hatırlatan aralıklı sesler çıkarması nedeniyle büyük ölçüde arttığı için tanıtıldı (Gottlieb, 1971, bölüm 7). Bu prosedürün görsel bir uyarana damgalamayı hızlandırdığı bilinmektedir (Smith ve Bird, 1963). 1. günde civcivler 45 dakika ("yetersiz" kontrol tavukları) veya 180 dakika ("aşırı eğitilmiş" deneysel tavuklar ) için uyarıldı . Daha sonra öğrenme sürecindeki yaklaşma tepkisinin kütlesine ve gücüne göre çiftler halinde gruplandırıldılar. Bu şekilde 10 çift tavuk seçilmiştir (bu çiftlerden birinde "yetersiz" kuş 60 dakika, "aşırı eğitilmiş" kuş ise 240 dakika uyarana maruz bırakılmıştır). Eğitimden sonra civcivler karanlık inkübatöre geri döndüler ve ertesi gün onlara radyoaktif urasil enjekte edildi. Daha sonra tavukların her biri dönen bir tekerleğe yerleştirildi ve 63 dakika boyunca bir uyarana maruz bırakıldı. Kullanılan uyaranın diğer deneylerde (Bateson, 1979) damgalamada etkili olduğu gösterildiğinden civcivler tercih testine tabi tutulmamıştır.
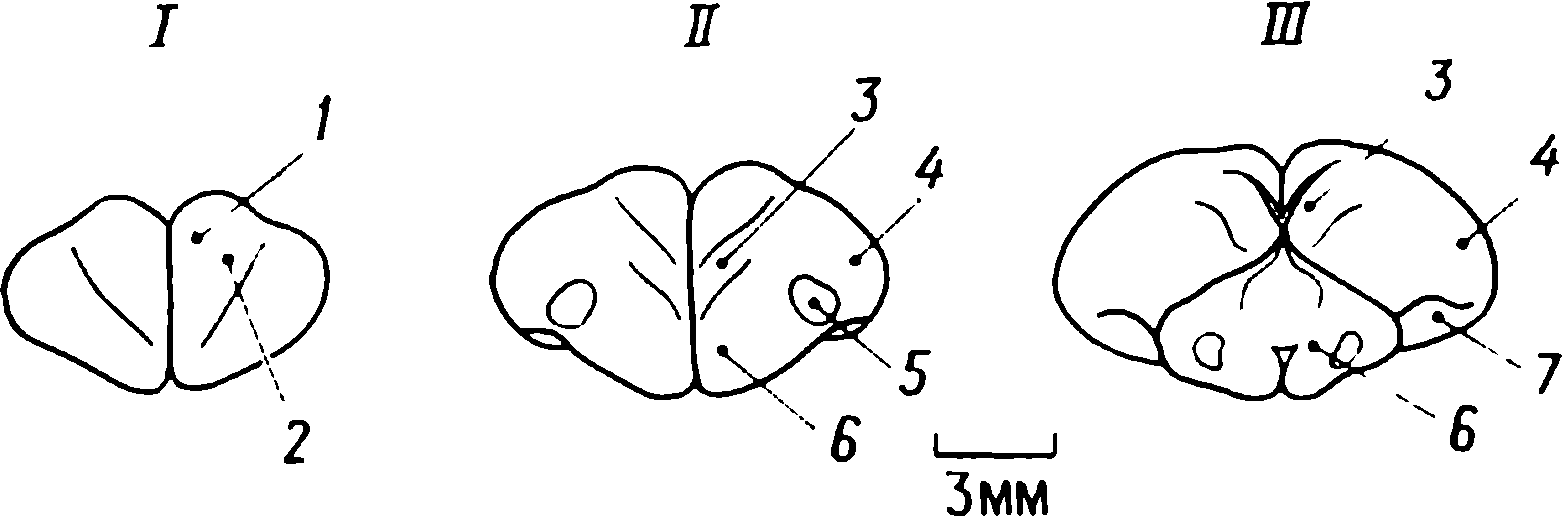
Enjeksiyondan 150 dakika sonra civcivler kurban edildi, beyinlerinin tamamı çıkarıldı ve katı karbondioksit ile donduruldu. Her beyin bir kaba yerleştirildi ve bir kod numarası verildi; sonraki tüm prosedürler kör bir şekilde gerçekleştirildi. Ön beyinden seri kesitler alındı, slaytlara yerleştirildi ve yukarıda açıklanan yöntem kullanılarak otoradyografi için hazırlandı. Kesitler daha sonra röntgen filmi ile kaplandı. Her bir tavuk çifti için pozlama aynı süre boyunca gerçekleştirildi, ardından film standart koşullar altında geliştirildi ve sabitlendi.
mikrodensitometre ile ölçüldü . Ölçümler için noktaların seçimi, teorik gerekçelerle ve ayrıca deneme deneyleri temelinde yapıldı. Onlarda, yukarıda açıklanan şekilde altı çift tavuk eğitildi. Bölümlerin imzaları görsel olarak incelendi ve her bir tavuk için öğrenmenin doğasını bilmeyen gözlemci, görsel analize dayalı öğrenme türü hakkında bir sonuca vardı. Bir çift tavukta fark yoktu. Kalan beş çiftten dördü için eğitimin doğası hakkında doğru sonuçlar elde edildi. Bu dört çift tavuğun hepsinde, ventral hiperstriatumun medial kısmı, özellikle serebral hemisferlerin ön ve arka kutupları arasındaki orta kısmı , aşırı eğitilmiş tavuklardan daha koyuydu (Şekil 4.2). Bir çift tavuktan alınan kesitler arasındaki başka hiçbir fark, öğrenilmiş davranıştaki farklılıklarla ilişkilendirilmedi. Öğrenmenin doğasını belirlemek için ventral hiperstriatum analizinin sonuçları, istatistiksel olarak anlamlı olmasa da , beynin bu bölümünün belirli bir önemini gösterdi. Bu nedenle ana deneyde ventral hiperstriatumun medial kısmının optik yoğunluğu ölçüldü. Ek olarak, hepsi görsel projeksiyon alanına ait olduklarından, aksesuar hiperstriatumda, orta ve dorsal hiperstriatum bölgesinde ve ektostryatumda yoğunluk ölçüldü (Şekil 4.2 ve A.5). Neostriatum ve memeli neokorteksine benzer olduğu düşünülen ön beyin bölgesinde de ölçümler yapıldı (Cohen ve Karten, 1974). Ayrıca, bir çekirdek kompleksi olan (Zeier ve Karten, 1971) arşistriatumun optik yoğunluğunu da ölçtük ; ancak, kompleksin ayrı ayrı bileşenlerini ayırt etmek mümkün değildi. Son olarak, ön beyin tabanı içinde, yarımkürenin ventromedial kısmında yer alan alanın optik yoğunluğunu ölçtük (Şekil 2.5, L).
, seri bölümlerin 18 radyootografına karşılık gelen yaklaşık 0.6 mm kalınlığında bir veya daha fazla "sütun" a bölündü. Farklı kuşların beyinleri işlenirken her yapı için bu tür sütunların sayısı sabitti. Her sütunda 10 radyootograf için elde edilen değerlerin ortalaması alınmış ve elde edilen ortalama değerler, belirli bir beyindeki tüm ölçümler için ortalama değerin yüzdesi olarak ifade edilerek standardize edilmiştir. Standardizasyon prosedürü, örneğin uygulanan radyoaktif urasil miktarındaki farklılıklara bağlı olabilecek farklılıkları ortadan kaldırdı. Ancak tüm ölçümler tamamlandıktan sonra beyni işaretleyen kod ortaya çıktı.
Standartlaştırılmış ortalama optik yoğunluk, iki bitişik sütuna karşılık gelen bir uzunluğa sahip olan ventral hiperstriatumun medial kısmında, yetersiz eğitilmiş tavuklarda aşırı eğitilmiş tavuklardan önemli ölçüde daha yüksekti . Bu sütunların yaklaşık konumu Şekil 1'de gösterilmektedir. 4.3, A. Bu bölgenin imza örnekleri Şek. 4.4. Diğer sütunlarda, "az eğitilmiş" ve "fazla eğitilmiş " tavuklar arasında hiçbir fark bulunmadı . Bununla birlikte, bazı sütunlardaki ölçüm sonuçlarının bir analizi, "yetersiz eğitilmiş" tavuklardaki bireysel göstergelerin dağılımının "aşırı eğitilmiş" tavuklardan daha fazla olduğunu göstermiştir. Kantitatif test için, varyasyon katsayısı, az eğitilmiş civcivlerdeki dağılım, aşırı eğitilmiş civcivlerdeki dağılıma bölünerek hesaplandı . Bu katsayılar, Şek. 4.3, B. Sütun 3 ve 4'te, "yetersiz eğitilmiş" tavuklar için göstergelerdeki varyasyonlar, "aşırı eğitilmiş" tavuklara göre önemli ölçüde daha yüksekti. Bu sütunlar, iki kuş grubu için ortalama değerlerde önemli farklılıkların bulunduğu sütunların hemen önünde yer alır (Şekil 4.3, E).
Bu çalışmanın sonuçları , ventral hiperstriatumun medial kısmında (5 ve 6 numaralı çubuklar)
Resim: 4.3. Ventral hiperstrinatumun medial kısmındaki seçilmiş alanların standardize edilmiş optik yoğunluğundaki ortalama farklar (“az öğrenilmiş” eksi “fazla öğrenilmiş”) . Seçilen bölgelerin yaklaşık yerleşimi, yukarıda gösterilen civciv beyni diyagramında, sırt yüzeyi çıkarılmış olarak gösterilmektedir (**P<0.01; ***P<0.001). B. Medial ventral hiperstriatum içindeki yedi bölümün her biri için standartlaştırılmış puanlardan hesaplanan dağılım oranları. Ordinat, "yetersiz eğitilmiş" tavuklardan elde edilen verilerin varyansının "aşırı eğitilmiş" tavuklardan elde edilen verilerin varyansına bölünmesiyle gösterilir (*P<0.05). (Noth ve diğerleri, 1979.)
0.50

Resim: 4.4. Bir çiftten "aşırı eğitilmiş" ve "yetersiz eğitilmiş" tavukların beyin bölümlerinin radyo-otografları. Fonksiyonel aktivite, belirli bir alanda bulunan radyoaktif urasil miktarı ile ölçülür. Yüksek kapsama bölgeleri, düşük dahil olma bölgelerinden daha koyudur; optik yoğunluk ne kadar yüksek olursa, inklüzyon da o kadar yüksek olur. "Aşırı eğitilmiş " bir kuşun imzası (B) , genellikle yetersiz eğitilmiş bir kuşun imzasından (A) biraz daha hafiftir . "Aşırı eğitilmiş" tavuğun beyninin tüm alanlarında daha düşük bir radyoaktif urasil dahil edildiğini gösteren bu etki, tüm tavuk çiftlerinde tutarlı değildi. Fark, eklenen haberci miktarındaki küçük farklılıklar gibi çeşitli faktörlere bağlı olabilir . Bu tür varyasyonlar, bir standardizasyon prosedürü kullanılarak ortadan kaldırılmıştır. Ventral hiperstriatumun medial kısmı ( her iki imzadaki üst oklar), yetersiz eğitim almış bir civcivin (A) radyo otografındaki çevreleyen alanlardan nispeten daha koyu ve aşırı eğitim almış bir civcivinkinden (B) önemli ölçüde daha koyu. Alttaki oklar (optik yoğunluklar sırasıyla 0.17 ve 0.15) , ön beyin tabanında ölçümler için seçilen alanı gösterir. Optik yoğunluk , ok uçlarının solundaki küçük bölgelerde ölçülmüştür .
değişiklikler hemen hemen her zaman meydana gelir, daha sonra diğer kısımlarında (3. ve 4. sütunlar) kaymalar yalnızca bazı kişilerde görülür . Bu çubuklar yaklaşık 2,4 mm uzunluğunda bir alanı kaplamaktadır. Başlangıçta MVG olarak adlandırdık (Hot ve diğerleri, 1979) ve bu kısaltma bazı araştırmacılar tarafından hala kullanılmaktadır ( Kossut ve Rose, 1984; Stewart ve diğerleri, 1984; Takamatsu ve Tsukada, 1985). Ancak bu alan medial ventral hiperstriatumun orta kısmını kaplar ve bu nedenle onu MVH olarak adlandırdık (Horn, 1981).
Başka hiçbir anatomik bölgede, iki tavuk grubu arasında önemli farklılıklar bulunmadı. Otoradyografi bulguları, önceki damgalama verileriyle iki önemli açıdan tutarlıdır. İlk olarak, PMCH, damgalamaya katılımı iki bağımsız deney serisiyle gösterilen (Bölüm 3.2 ve 3.3) serebral hemisferlerin bir parçası olan önbeyin çatısının ral kısmının büyümesi içinde yer alır. İkincisi, değişimin yönü yukarıda açıklanan çalışmadakiyle aynıydı (Bölüm 3.2), yani yumurtadan çıktıktan sonraki 1.
, habercinin dahil edilmesindeki değişikliklerin bir şekilde damgalama süreciyle ilgili olduğunu düşündürür . Bununla birlikte, otoradyografi verilerinin başka bir yorumu da mümkündür. Bu deneyde, görsel bir uyaranın sunumuna evcil bir tavuğun annelik çağrısı eşlik etti . Sonuç olarak, 1. günde "yeniden eğitilen" kuşlar daha uzun süre işitsel uyarı aldı. İşitsel projeksiyon bölgesi - L alanı - neostriatumun arka kısmında, ventral hiperstriatumun altında ve yanında yer alır (Şekil A.6). Her iki yapı da belirgin bir lifli tabaka ile ayrılır. Ventral hiperstriatumun medial kısmının optik yoğunluğu, L alanının dışında bu katmanın üstünde ve medialinde ölçülmüştür.Bununla birlikte, beç tavuğu (Numidia meleagris) ve evcil tavukta, L alanı ventral hyperstriatum (Bonke et) ile ilişkilidir. ark., 1979; Bradley ve ark., 1985), böylece bu yapının hücreleri işitsel uyarımdan etkilenebilir. Ventral hiperstriatumun en kaudal kısmı bile L alanından projeksiyonlar alır. Bununla birlikte, ataların bu kısma dahil edilmesi, aşırı eğitilmiş ve yetersiz eğitilmiş kuşların farklı davranışsal deneyimlerine bağlı olarak değişmedi (Şekil 4.3, çubuk 7). Bu sonuçlar, "işitsel deneyim"in anlamı hakkındaki hipotezleri desteklememektedir; ne de bölünmüş beyinli tavuklar üzerindeki çalışmaların sonuçları (Bölüm 3.1). Bu çalışmada önbeyin çatı bölgesinde radyoaktif urasil'in RNA'ya dahil edilmesinde interhemisferik bir asimetri gözlendi.Bu interhemisferik asimetri, civciv eğitim sırasında her iki kulağı da kullandığından işitsel uyarımdaki bir farkla açıklanamaz.
Radyoaktif 2-deoksiglukoz kullanarak araştırma
Yukarıda açıklanan imza verilerinin yayınlanmasından kısa bir süre sonra, Kozaka ve diğerleri (Kohsaka ve diğerleri, 1979), tavuklarda damgalama üzerine kendi çalışmalarını yayınladılar. Ayrıca radyootografik analiz kullandılar, ancak radyoaktif urasil değil, 2-deoksiglukoz kullandılar. Bu yöntem ilk olarak Sokolov ve arkadaşları tarafından önerilmiştir (Sokoloff ve diğerleri, 1977). Çoğu koşulda nöronların glikozu emdiği ve onu bir enerji kaynağı olarak kullandığı gerçeğine dayanır . Bir nöron aktif olduğunda, diğer şeylerin yanı sıra , akson terminallerindeki iyon konsantrasyonunu ve nörotransmiterlerin sentezini eski haline getirmek ve sürdürmek için enerjiye ihtiyaç duyar. Bu enerji gereksinimi glikoz tarafından karşılanır. İkinci karbon atomunda bir oksijen atomu olmayan glikoz analoğu - 2-deoksiglikoz, hücre tarafından sıradan glikozla aynı şekilde emilir. Ancak ikincisinden farklı olarak 2-deoksiglikoz zayıf bir şekilde parçalanır ve hücrede kalır (Sokoloff ve diğerleri, 1977; ayrıca bkz. Hawkins ve Miller, 1978). Sokolov ve diğerleri, 2-deoksiglikozu 14 C ile etiketlediler. Bu radyoaktif madde farelere verildiğinde , beyin bölümlerinin imzalarını hazırlayarak nöronların artan aktivitesinin olduğu bölgeleri ortaya çıkarmak mümkün oldu . Sokolov yöntemi, beynin nöronların dürtü aktivitesinin enerji maliyetlerinin anında yenilenmesini gerektirdiği alanlarını belirlemek için kullanılabilir. Radyoaktif urasil kullanan yöntem , başta RNA olmak üzere makromoleküllerin sentezinde ve metabolizmasında değişikliklerin meydana geldiği alanların belirlenmesini mümkün kılar . Bu değişikliklerin, nöronların dürtü aktivitesinin doğrudan sonuçlarını yansıtması olası değildir ; bunun yerine, bu yöntem, aktivitenin nörosekresyon veya büyüme gibi dolaylı etkilerini ortaya çıkarır . Bu iki yöntem, nöronal metabolizmanın farklı yönlerini ortaya koysa da, bulunan yönler tamamlayıcı olabilir .
Kozaka ve diğerleri (Kohsaka ve diğerleri, 1979) civcivleri yumurtadan çıktıkları andan 8-12 saat 45 dakikaya kadar karanlıkta büyüttüler. Bundan sonra, tavuklar iki gün boyunca bireysel karanlık kafeslere geri gönderildi. Üçüncü gün, her bir civciv 45 dakika boyunca kırmızı topla birlikte tekrar yastığa yerleştirildi. Hareket eden cismi 20 dakika boyunca takip eden civcivlerin topun üzerine basıldığı ve 5 dakikadan daha az takip eden civcivlerin damgalanmadığı kabul edildi. Başına-
civcivler daha sonra bir gün daha karanlık inkübatöre geri gönderildi. Yaşamın dördüncü gününde civcivlere radyoaktif 2-deoksiglukoz enjekte edildi ve yine 45 dakika boyunca hareket eden kırmızı bir top sunuldu . Bu prosedürün amacı, görsel analizördeki ve damgalama meydana geldiğinde oluşabilen sinir ağlarındaki aktiviteyi harekete geçirmekti. Uyarılmış nöronların radyoaktif molekülleri emmesi gerekiyordu , böylece radyootograflar bu hücresel sistemlerin lokalizasyonunu belirleyebiliyordu. Bunun için beyinden kesitler yapıldı ve röntgen filmine yerleştirildi.
Kozaka ve meslektaşları, damgalanmış tavukların, damgalanmamış kontrollere kıyasla önemli ölçüde daha yüksek medial ventral hiperstriatal inklüzyona sahip olduğunu bildirdi. Ventral hiperstriatumun aktif kısmı , serebral hemisferlerin ön ve arka kutuplarını birleştiren çizginin yaklaşık olarak ortasına yerleştirildi ve bu nedenle PMSH'ye karşılık geldi. Aynı bölümler , damgalanmış kuşların neostriatumunun yan kısmında artan bir dahil olma seviyesi de gösterdi. Serebral hemisferlerin lateral kısmının damgalama sürecindeki olası rolü aşağıda tartışılmıştır (s. 118-119). İki tavuk grubu arasında başka hiçbir önemli bölgesel farklılık bulunmadı.
Kozaki ve meslektaşlarının deneylerinin yorumu tam olarak net değil. Örneğin yazarlar, yukarıda açıklanan yöntemle eğitilen tavukların tanıdık bir uyaran için bir tercih geliştirip geliştirmediğini belirtmezler. Civcivlere kırmızı bir balon ile daha önce görmedikleri başka bir nesne arasında seçim hakkı verilmedi. Tarif edilen deneylerde veya başka bir deneyde böyle bir test yapılmadıysa, hareket eden topu takip eden civcivlerin topun üzerine baskı yaptığından emin olunamaz. Nesneyi takip eden kuşların , kırmızı topun takip edecek kadar çekici geldiği aktif bireyler olması muhtemeldir. Bununla birlikte, bu sonuçlar bir dereceye kadar ihtiyatla kabul edilirse, bir kez daha damgalamanın ortaya çıkmasının, beyin aktivitesi ve davranışlarının çok farklı göstergelerine dayanarak değerlendirilebilen PMVP'de değişikliklere yol açtığı gerçeğine tanıklık ederler.
ventral hiperstriatumun medial kısmına artan bir 14C - 2 -deoksiglukoz dahil edildiğini bulmuşlardır . Bu alan görünüşe göre PMSH'nin en rostral kısmıyla örtüşüyor. Ventral hyperstriatum'un lateral kısmında da artmış inklüzyon bulundu ve neo-
140 striatum ve ek hiperstriatum. Bu deneylerde, civcivler doğumdan kısa bir süre sonra 2 saat boyunca 1.8 veya 2.5 kHz'lik aralıklı tonlara maruz bırakıldılar.Tercih bir Y labirentinde test edildi. Her dalın ucuna kutular yerleştirildi. Her birinin arkasında bulunan hoparlörler 1.8 ve 2.5 kHz'lik tonlar yayıyordu. Y'nin tabanına bir civciv yerleştirildi ve sesin orijinal olarak verildiği dala yaklaşma kriterine ulaşırsa bir iz geliştirdiği kabul edildi . Üçte üçü veya dörtte üçü doğru branş seçimi kriter olarak alınmıştır. Tercih, 1,8 kHz ton ile eğitilen 11 civciv (Grup 1) ve 2,5 kHz ton ile eğitilen 4 civciv (Grup II) üzerinde test edildi . Grup I'de 8 civciv doğru seçimi yaptı ve 3 civciv yapamadı. Grup II'de 3 civciv doğru tercih yapmış; birinin tercihi yoktu. Bu sonuçlar rastgeleden önemli ölçüde farklı değildir. Bu nedenle , tavukların imprinting geliştirdiğini gösteren eksiksiz bir veri yoktur ve bu nedenle, "eğitimli" ve "eğitimsiz " beyindeki dahil olma farklılıklarının biyolojik anlamı belirsizliğini koruyor.
Mayer ve Shaikh'in çalışmasının şaşırtıcı bir detayı, tanımladıkları görsel Wulst'a işaretli 2 -deoksiglikoz eklenmesindeki artıştır. Kontrol tavukları eğitilmedi. 14 C-2-deoksiglukoz enjekte edildi ve 1.8 kHz'lik bir tona maruz bırakıldı ve ardından puanlandı. "Eğitilmiş" kuşlarda ek hiperstriatuma dahil olma, kontrol grubundan daha yüksekti . Çalışmada kontrol tavuklarının 1.8 kHz'lik bir tona maruz kalmadan önceki görsel deneyimi anlatılmamıştır. Ve bu bilgi önemlidir: görsel projeksiyon alanına daha yüksek düzeyde dahil olma, akustik baskıya değil, "eğitimli" kuşların ve kontrol grubunun farklı görsel deneyimlerine bağlı olabilir.
Mayer ve Sheikh'in çalışmasının sonuçları , PMVP'nin bir kısmının beç tavuğu civcivlerinde işitsel baskıda ve aynı zamanda evcil tavuklarda görsel baskıda olma olasılığını tahmin ettikleri için caziptir. Bu nedenle, bu çalışmada ortaya çıkan belirsizliklerin ortadan kaldırılması acildir.
Bazı çözülmemiş sorunlar
Radyootografik çalışmalar , evcil tavuklarda imprinting ile ilişkili biyokimyasal değişikliklerin , ön beyin çatısının yan kısmının tüm büyümesinin özelliği olup olmadığı veya bazı bölgelerinin özelliği olup olmadığı sorusunun cevabına yaklaşmayı mümkün kılar . Bununla birlikte, yerel kaymaların saptanması, beynin ölçümlerin yapılmadığı veya incelediğimizden farklı bir gelişim dinamiklerine sahip olduğu alanlarda değişiklik olasılığını ortadan kaldırmaz ( bkz. Bölüm 7) . Ek olarak, yerel ve yaygın kaymaların aynı anda var olması mantıksal olarak mümkündür, ancak ikincisi, yöntemin belirli bir hassasiyeti için "gürültü seviyesinin altında" çıkıyor. Bu olasılıkların akılda tutulması gerektiği açıktır .
Yerel değişikliklerin saptanması, kişinin birçok soruyu gündeme getirmesine ve damgalamanın daha fazla araştırılması için çok çeşitli yöntemler uygulamasına olanak tanır. Kullanılan radyootografik yöntem, hücresel düzeyde lokalizasyonu belirlememize izin vermez. Birleşmenin esas olarak PMIG'de bulunan hücrelerin gövdelerinde meydana gelmesi mümkündür, çünkü amino asitlerin toplandığı ana ribozom kütlesinin bulunduğu gövdelerdedir. Urasil birleşme seviyesindeki bir artış paralel olarak ortaya çıkarsa veya nöronlardaki yapısal değişikliklere eşlik ederse, bu değişikliklerin nerede bulunabileceğini bilmek önemlidir ; örneğin, tamamen SMVG alanıyla sınırlı olan dendritlerin ve aksonların arborizasyonlarında mı yoksa bu alanın ötesine uzanan akson terminallerinde mi meydana geldiği . Bilginin PMCH'ye girme ve çıkma yollarını, PMCH'de sinyallerin ne tür fizyolojik dönüşümünün meydana geldiğini ve sito-mimarisinin, nörotransmitterlerinin neler olduğunu, bu alanın uyarılmasının veya çıkarılmasının davranış üzerindeki etkilerini, etkilerini bilmek önemlidir. Beynin bağlantılı olduğu diğer alanlarında vb. Soruların listesi neredeyse sonsuzdur ve damgalamanın hücresel temelini anlamak için birçoğunun yanıtlanması gerekir.
Özet
Baskı işlemi için önemli olan ön beyin çatısının rostral kısmı (Bölüm 3) nispeten büyük bir yapıdır. Baskı oluşumunda tüm yapının mı yoksa sadece sınırlı bir kısmının mı yer aldığı sorusunu aydınlatmak için radyootografik yöntem kullanıldı. Bir çalışma, etiketli haberci olarak radyoaktif urasil kullandı. Öğrenme ile ilişkili dahil edilmesindeki değişiklikler , ön beyin çatısının yalnızca sınırlı bir bölümünde, ventral hiperstriatumda bulundu . Bu yapının sadece orta hatta yakın bir bölümünde belirgin bir inklüzyon kayması gözlendi. PMCH olarak adlandırdığımız bu alan yaklaşık 2,4 mm uzunluğundadır ve yaklaşık olarak serebral hemisferlerin ön ve arka kutuplarının ortasında yer alır. Etiket olarak 14C-2-deoksiglukoz kullanıldığında evcil tavukların beyninde benzer değişiklikler bulundu .
7*
EDEBİYAT
Bateson P. Civcivlerde damgalama sırasında yeni bir uyarana kısa süre maruz kalma ve bunun sonraki tercihler üzerindeki etkisi, Anim. Leam. Behav., 1979, 7, 259-262.
Bonke BA, Bonke D., Scheich H. Gine Tavuğundaki (Numida meleagris) işitsel ön beyin çekirdeklerinin bağlantısı , Celi Tissue Res., 1979, 200, 101-121.
Bradley P., Davies DC, Horn G. Evcil civcivde (Gallus domesticus) hyperstriatum ventrale'nin bağlantıları , J. Anat., 1985, 140, 577-589.
Cohen DH, Karten HJ Kuş beyninin yapısal organizasyonu: genel bir bakış. İçinde: Kuşlar, beyin ve davranış (editörler 1. J. Goodman ve M.W. Shein), 1974, s. 29-73. Akademik Basın, New York.
Gottlieb G. Kuşlarda tür tanımlamasının gelişimi: algının doğum öncesi belirleyicilerine yönelik bir araştırma. Chicago University Press, Chicago, 1971. Hawkins RA, Miller AL Bilinçli farelerin beyinlerinden radyoaktif 2-deoksi-D-glikoz-6-fosfat kaybı: bölgesel glikoz kullanımının kantitatif otoradyografik tayini için çıkarımlar, Neuroscience, 1978, 3, 251 -258.
Horn G. Sinirsel öğrenme mekanizmaları: evcil civcivde damgalamanın analizi, Proc. R. Soc., 1981, B213, 101-137.
Horn G., McCabe BJ Beyinde asitte çözünmeyen bileşiklere urasilin dahil edilmesini incelemek için bir otoradyografik yöntem, J. Physiol., Londra., 1978, 275, 2-3P.
Horn G., McCabe BJ, Bateson PPG Damgalamadan sonra civciv beyninin otoradyografik bir çalışması, Brain Res., 1979, 168, 361-373.
Kohsaka S., Takamatsu K-, Aoki E., Tsukada W. Civciv beyninin [14C] 2-deoksiglukoz tekniği kullanılarak imprinting sonrası metabolik haritalaması , Brain Res., 1979, 172, 539-544.
Kossut M., Rosse SPR Pasif kaçınma eğitimi sırasında civciv beyin yapılarına Diferansiyel 2-deoksiglikoz alımı. Nörobilim, 1984, 12, 971-977.
Lashley KS Beyin mekanizmaları ve zekası: beyindeki yaralanmaların niceliksel bir çalışması . Chicago University Press, Şikago, 1929.
Lashley KS Engram arayışında, Symp. sos. tecrübe. Biol., 1950, 4, 454-482.
Maier V., Scheich H. Akustik baskı, civciv ön beyninde diferansiyel 2-deoksi-D-glikoz alımına yol açar, Proc. nat. Acad. bilim ABD, 1983, 80, 3860-3864.
Rogers AW Otoradyografi Teknikleri (2. baskı). Elsevier, Amsterdam, 1973,
Smith FV, Bird MG Farklı duyusal modalitelerdeki uyaran kombinasyonlarının evcil civciv için göreli çekiciliği, Anim. Behav., 1963, 11, 300-305.
Sokoloff L., Reivich M., Kennedy C., Rosiers MHD, Patlak C. S. Pettigrown KD, Sakurada O., Shinohara M. Lokal serebral glikoz kullanımının ölçümü için 14 C-deoksiglukoz yöntemi: teori, prosedür ve bilinçli ve anestezi uygulanmış albino sıçandaki normal değerler, J. Neurochem., 1977, 28, 897-916 .
Siewart MG, Rose SPR, King TS, Gabbot PL A „ Bourne R. Pasif kaçınma eğitiminin ardından civciv medial hyperstriatum ventrale'deki sinapsların hemisphe rio asimetrisi: stereolojik bir araştırma, Devi . Brain Res., 1984, 12, 261-269.
Takamatsu K., Tsukada U. Civciv ve ördek yavrusu damgalamanın nörobiyolojik temeli. In: Nörobilimlerde Perspektifler: molekülden akla. (ed Y. Tsu kada), 1985, s. 187-206, Springer, W. Berlin.
Zeier H., Karten HJ Güvercin arşivi: afferent ve efferent bağlantıların organizasyonu, Brain Res., 1971, 31, 313-336.
BÖLÜM 5
BEYİN HASARI, HAFIZA İZLERİNİN OLUŞUMU VE SAKLANMASI
Bölümde açıklanan deneyler. 3, orijinal biyokimyasal çalışmalarda ortaya atılan sorunun yanıtına yaklaşmamıza izin verir : özellikle imprinting ile ilişkili biyokimyasal değişiklikler var mı? Deneylerin gösterdiği gibi, baskılama için PMVP gerekliyse, PMVP hasar gördüğünde baskılama bozulmalıdır. Bu varsayım, PMVG'yi yok ederek ve ardından bir baskı geliştirmeye çalışarak test edilebilir. Böyle bir test sadeliği ile dikkat çekiyor, ancak bir takım dezavantajları var. Bir yandan, beyin gelişiminin erken bir aşamasında hasar görürse, beynin başka bir bölümünün hasarlı bölgenin işlevlerini üstlenmesi olasıdır. Bu durumda, deney beklenen sonuçlara yol açmazsa, bu, ilk varsayımın yanlışlığını değil, gelişen beynin plastisitesini gösterecektir. Öte yandan, beklenen sonuç elde edilirse, bu kesinlikle PMVP'nin baskılama için atlanmadığını gösterecektir, ancak bu bağımlılık oldukça önemsiz olabilir. Örneğin, PMVP'nin hasar görmesi civcivlerin daha az aktif, daha az uyanık, daha az motive veya görme engelli olmasına neden oluyorsa, bir damgalama uyaranı ile sunulduğunda zayıf öğrenecekler veya hiç öğrenemeyeceklerdir . Ek olarak, PMIG, tıpkı sıralı olarak organize edilmiş bir devredeki bu devrenin çalışması için korunması gereken herhangi bir bağlantı gibi, beynin diğer herhangi bir bölümüyle aynı şekilde damgalama için gerekli olabilir. Hasar yönteminin listelenen sınırlamaları çok önemlidir . Bununla birlikte, bu yöntemin damgalama çalışmasında kullanılmasına ilişkin argümanların yeterince ikna edici olduğunu düşündük.
PMVG hasarı ve tercih gelişimi
PMVP hasarının tercih üretimi üzerindeki etkisini araştırmayı amaçlayan deneylerde iki grup tavuk kullanıldı (McCabe ve diğerleri, 1981). Bir grup civciv
O 11 12 12,5 14
A I P -ı b 2 - ■ "
B I R — ii □ 1
Resim: 5.1. Damgalama sırasında tercihin geliştirilmesinde PMVC'ye verilen iki taraflı hasarın etkisini analiz etmek için bir deneydeki prosedürler dizisinin şeması (McCabe ve diğerleri, 1981). Üst satırın üzerindeki sayılar, yumurtadan çıktıktan sonraki saat cinsinden (0 saat) çeşitli maruziyetlerin yaklaşık süresini gösterir. Belirtilen süre , alt satırda gösterilen ilgili olaylar için de geçerlidir . Kesintisiz çizgi ile gösterilen süre boyunca civcivler karanlık inkübatörün izole bölmelerinde tutuldu. Civcivler yaklaşık 47 saatlik olduklarında tercih testi yapıldı. Tüm tavuklara, 1. günde baskı nesnesine maruz bırakılmadan önce 30 dakika boyunca ön uyarı (P) verildi. Prosedürlerin sırası : A. PMVG'si hasar görmüş civcivler (hasar siyah karelerle işaretlenmiştir). Satır sonu ile işaretlenen süre boyunca, civcivler damgalama uyaranına maruz bırakıldı. B. Sham kontrollü kontrol tavukları. Beyaz dikdörtgenle gösterilen süre boyunca civcivlere anestezi uygulandı, kafa derisine kesikler atıldı ve dikildi. Tüm semboller diğer benzer şekillerde kullanılmaktadır.
Lyat, PMVP'ye zarar verdi, ikincisi kontrol görevi gördü. Yumurtadan çıktıkları andan itibaren civcivler karanlıktaydı, ardından 30 dakika boyunca tepeden aydınlatmaya maruz bırakıldılar, ardından 30 dakika boyunca baskı nesnesinin önünde dönen bir tekerleğe yerleştirildiler (Şekil 5.1). PMVD yaralanmasından önce her iki civciv grubuna damgalama nesnesinin bu ilk sunumu, yalnızca aktif kuşları seçmek ve böylece bu deneyde yetersiz öğrenen civcivlerin sayısını en aza indirmek için yapıldı (bkz. s. 89). Civcivler, 30 dakikalık aktivitelerine göre çiftler halinde eşleştirildi (Şekil 5.2, eğitim süresi /). Her çiftten bir tavuk deney grubuna, diğeri kontrol grubuna alındı. Seçim prosedürünün potansiyel bir dezavantajı, bu 30 dakika boyunca tercih oluşumunun meydana gelme olasılığıydı. Bununla birlikte, bir deneme deneyinde, 30 dakika boyunca bu şekilde eğitilen tavukların, ertesi gün tanıdık ve yeni uyaranlar arasında seçim yapma durumunda herhangi bir tercih göstermediğini bulduk.
Civcivlerin yarısına, kırmızıya boyanmış iki büyük yüzeyi olan ve dakikada 28 devir frekansında dönen bir kutu (Şekil 2.3, H) verildi. Diğer yarısına farklı bir uyaran sunuldu (Şekil 2.3, C) - evcil tavuğun atası olduğu düşünülen bir tür olan banka tavuğunun (Gallus gallus) sallanan büstü (Zeuner, 1963). Maruz kalma sırasında, tavuğun annesinin çağrısının manyetik bir kaydı duyuldu. Her çiftin (deney ve kontrol grupları) her iki tavuğuna da aynı damgalama nesnesi sunuldu.
Resim: 5.2. Deney civcivleri ve kontrol civcivleri için dört eğitim periyodu boyunca yaklaşma tepki puanları . Yaklaşım göstergeleri, saatteki tekerlek devir sayısı olarak ifade edilir, ortalama değerler verilir . Sahte çalıştırılan kuşlar için (beyaz çubuklar), dönem II'deki öğrenme oranları, dönem I'dekinden önemli ölçüde daha yüksektir (Wilcoxon istatistikleri, T=3\ P<0.01) ve dönem III'te , dönem II'dekinden daha yüksektir (î = 5) ; P<0.01). Bu grubun III . ve IV . dönemlerdeki göstergeleri önemli ölçüde farklılık göstermemektedir. Beyin hasarlı civcivlerde (taranmış parmaklıkların arkasında) 2. gündeki yaklaşma tepkisi, her üç dönem boyunca da kontrollerden önemli ölçüde daha düşüktür. (McCabe ve diğerleri, 1981.)
Bir çift tavuğu 30 dakika eğittikten sonra anestezi uygulandı . Bunlardan birinde kafatasına küçük bir kesi yapıldı ve ucu yalıtımsız bir iğne PMVP'ye yerleştirildi. Yüksek frekanslı bir akım geçtiğinde, iğnenin ucunun etrafındaki doku pıhtılaştı. Her iki tarafta PMVG pıhtılaşması yapıldı. Ameliyattan sonra saçlı deri dikildi. Kontrol tavuklarında baş derisi aynı şekilde kesildi ve dikildi . Bundan sonra, anestezi altındaki her iki kuş da ayırt edilemez hale geldi. Her birine bir kod numarası verildi ve daha ileri eğitim ve test prosedürleri , deneyi yapan kişinin hangi kuşun beyin hasarı aldığını bilmeyeceği şekilde gerçekleştirildi.
Ameliyatın ertesi günü anestezinin etkisi geçince 2,5 saat daha aynı uyaran kullanılarak civcivlerin eğitimine devam edildi. Bundan sonra (Şekil 5.1), damgalama oluşumu sırasında hangi uyaranın sunulduğuna bağlı olarak hem tanıdık hem de yeni uyaranlara, kırmızı bir kutuya veya doldurulmuş bir banka tavuğuna yaklaşımlarını karşılaştırarak tüm tavukların tercih tepkisi ölçüldü . Test , önceki deneylerde kullanılandan farklıydı . Dönen bir tekerleğe yerleştirilmiş tavuğun 4 dakika boyunca farklı sekanslarda tanıdık ve yeni nesnelerin gösterilmesinden oluşuyordu. Tercih endeksi hesaplandı, yani yüzde olarak ifade edilen , tanıdık bir nesneye yöneltilen tüm yaklaşım tepkilerinin payı; elde edilen değer, yeni uyarana göre bu uyaranın çekiciliğinin bir değerlendirmesini verdi. Gösterge aşağıdaki formüle göre hesaplanmıştır :
10X (Bilinen uyarana
yaklaşma tepkilerinin sayısı) (Bilindik uyarana yaklaşma tepkilerinin sayısı)-{-(
Yeni uyarana yaklaşma tepkilerinin sayısı)
Tercih puanı sıfır (yalnızca yeni bir uyarana yaklaşma) ile 50 (tercih yok) ve 100 (yalnızca tanıdık bir uyarana yaklaşma) arasında değişiyordu.
Eğitim prosedürü sırasında, sahte çalıştırılan kontrol civcivlerinin yaklaşma tepkisi arttı ve ardından belirli bir seviyede stabilize oldu (Şekil 5.2). Tekerleğin 1 saatteki devir sayısı olarak ifade edilen yaklaşıklık, / döneminde zayıftı . 2. gün antrenmanın 1. saatinde yaklaşma tepkisi arttı, sonraki saatte artmaya devam etti ve sonra sabitlendi. Deney civcivlerinde I periyodundaki yaklaşım puanları kontrol grubu ile hemen hemen aynıydı. Bu, deneysel ve kontrol kuşlarının ameliyat öncesi eğitim döneminde yaklaşma etkinliği düzeyi açısından iyi bir şekilde eşleştiğini göstermektedir . Ancak, sadece bu dönemde her iki grup için yakınlaşma oranları benzerdi. Beyin hasarlı tavukların ortalama yaklaşma puanları zamanla artmadı ve sürekli olarak kontrol grubunun puanlarının altındaydı. Bu öğrenme puanı hesaplanırken , diğerlerinde olduğu gibi, kırmızı kutuya maruz bırakılan tavuklardan elde edilen veriler, doldurulmuş tavuğa maruz bırakılmadan elde edilen verilerle toplandı, çünkü ilgili analiz, kullanılan nesnenin doğasına bağlı olarak ortalama puanlarda farklılık göstermedi. öğrenirken.
İki tavuk grubu için tercih puanları şekil 2'de gösterilmektedir. 5.3. Sahte çalıştırılan kontrol tavukları, açıkça tanıdık nesneyi tercih ettiler. Beyin hasarlı kanatlılarda böyle bir tercih yoktu ve ortalama puanlar 50'den önemli ölçüde farklı değildi.
Tercih puanlarını hesaplamak için kullanılan denklemden, üç şekilde azaltılabileceği görülebilir: 1) tanıdık bir nesneye olan yakınlığı azaltarak; 2) yeni bir nesneye yaklaşımı arttırmak ; 3) 1 ve 2'nin bir kombinasyonu ile. Deneyselin ortalama yaklaşımı ve

Resim: 5.3. Yaşamın ikinci gününde 2,5 saat boyunca bir damgalama uyaranına maruz kaldıktan sonra PMVG'ye iki taraflı hasar veren ve sahte ameliyat edilen tavuklarda (LS) tercih indeksleri. Deney civcivlerinde ortalama tercih indeksi %56.58±9.67'dir. Bu gösterge, %50'den önemli ölçüde farklı değildir, ancak sahte olarak çalıştırılan kontrol grubu için göstergeden önemli ölçüde farklıdır (7=2.93; P<0.02). Tercih göstergesinin y ekseni boyunca çizildiği diğer tüm şekillerde olduğu gibi bu şekilde de kesikli çizgi tercihin yokluğuna karşılık gelen seviyeyi gösterir. (McCabe ve diğerleri, 1981.)
Sahte ameliyat edilen tavuk grupları tabloda verilmiştir. 5.1. Sahte ameliyat edilen tavuklar, test durumunda yeni nesneye tanıdık uyarıcıdan çok daha az yaklaştı. Buna karşılık, beyin hasarı olan tavuklarda tanıdık bir uyarana yaklaşmak, yeni bir uyarana yaklaşmaktan önemli ölçüde farklı değildi . Bu nedenle, bu gruptaki daha düşük yaklaşma puanları, esas olarak tanıdık bir nesneye yaklaşma tepkisindeki azalmaya bağlıydı. Deney civcivleri, ona kontrol civcivlerinin yeni nesneye davrandıkları gibi davrandılar . Bu sonuçlar, beyin hasarlı tavukların kendilerine sunulan uyaran hakkında hiçbir şey hatırlamadıklarını göstermektedir .
Bu deneyin sonuçları farklı şekillerde açıklanabilir (örneğin bkz. s. 101). Bununla birlikte, PMVP'si hasar görmüş tavuklarda baskı yapma yeteneğinin bozulması, aşağıdakilere bağlı olamaz:
Tablo 5.1. Bir tercih testinde yaklaşıklık; tanıdık ve yeni nesnelere doğru ortalama tekerlek dönüş sayısını gösterir (McCabe ve diğerleri, 1981'den sonra)
nesneye yönelik toplam etkinliğin yüzdesini temsil ettiğinden, genel etkinlikte bir azalma . Ayrıca beyin hasarlı civcivlerin yeni bir cisme yaklaşımı kontrol grubununkine benzerdi (Tablo 5.1). Eğitim ve test sırasındaki yaklaşma tepkisinin bozulması, beyin hasarlı civcivlerde herhangi bir duyusal veya motor bozukluğa mı bağlıydı? Bu soruyu cevaplamak için , bu fonksiyonlardaki büyük değişiklikleri ortaya çıkarabilecek bir test kullanıldı . Ucu zayıf ışıklı bir çubuk, karanlık bir inkübatöre yerleştirildi ve tavuktan 30 cm uzaklıkta bir yandan diğer yana hareket ettirildi. Civcivlerin çubuğa yaklaşması ve gagalamaya başlaması için geçen süre ölçülmüştür. Deney civcivlerinin harcadığı süre ( ortalama değer 39,5 sn) kontrol grubundaki süreden (45,5 sn) önemli ölçüde farklı değildi. Bu el-göz testi ameliyatlı ve sahte ameliyatlı kuşlar arasında hiçbir fark göstermedi . Son testte , deney tavuğu, sahte olarak çalıştırılan çiftiyle birlikte, üzerinde fırınlar, pipetler, aletler, kitaplar, kalemler vb. bulunan çeşitli nesnelerin bulunduğu bir laboratuvar masasına yerleştirildi. değerlendirildi - nesneleri gagalama, engellerden kaçınma ve genel fiziksel aktivite. Hasarlı MVP'ye sahip civciv, kontrol çiftinden ayırt edilemedi.
Hasarın yeri ve boyutu, beynin seri kesitlerinde değerlendirildi (Şekil 5.4). Her deney tavuğunun ventral hyperstriatum'un tahrip olmuş bir kısmı vardı. PMIG dahil olmak üzere hiperstriatum alanı IV ve VII dilimleri arasında yer alır. Bu alan tüm deney tavuklarında önemli ölçüde hasar görmüştür.

ben ben
3 mm
Resim: 5.4. Tavuk beyninin şematik kesitlerindeki lezyonların rekonstrüksiyonu. Dilimler arasındaki mesafe 500 mikrondur; en rostral dilim /'dir. Siyah, 12 civcivin tamamında yok edilen alanları gösterir . Tüm tavukların dörtte birinden fazlasında gölgeli alanlar yok edildi. Tanımlamalar: 1 — ek hiperstriatum; 2 - ara hiperstriatum; 3 - dorsal hiperstriatum ; 4 - ventral hiperstriatum; 5 - neostriatum; 6 - ektotriyum; 7 - koku alma payı; 8 - yükselen paleostriatum; 9 - mide kızları; 10 - ilkel paleostriatum; 11 - arşiv. (McCabe ve diğerleri, 1981.)
Civcivler uyarıcının özelliklerini tanımayı öğrendikçe uyarana yaklaşma etkinliği artar (Salzen ve Sluckin, 1959; Hom ve diğerleri, 1973). Bu nedenle, eğitim sırasında ameliyat edilen kuşların düşük aktivitesi, bozulmuş öğrenme yeteneğinin bir sonucu olabilir. Bu nedenle, eğitim ve test sırasında ortaya çıkan davranış farklılıkları, ameliyat edilen kuşlardaki bir kusurun , yani kendilerine sunulan uyaranın özelliklerini hatırlayamamalarının sonucu olabilir.
Test
O 21 22 25 28 47
4 • P1 ben ■■ 1
B i R —• ben —□ •
Resim: 5.5. PMVP'nin yok edilmesinin, damgalama sonucunda elde edilen bir tercihin izlerinin korunmasını nasıl etkilediğini açıklamak için tasarlanmış bir deneyin şeması. A. Beyni hasarlı kuşlar B. Yanlış çalıştırılan kontrol grubu.
PMSH'nin damgalama için gerekli olması durumunda, yok edilmesinin damgalamanın imkansızlığına yol açacağı varsayımlarını test etmek için beyin yıkımı ile deneyler yapıldı . Deneyin sonuçları bu varsayımı doğrular ve bu nedenle, damgalama sürecinde PMVP'nin belirli bir rolü olduğu hipotezini destekler. Bununla birlikte, yukarıda belirtildiği gibi (s. 101), beyin yok eden deneylerden elde edilen verileri yorumlarken dikkatli olunması gerekir ve bizim sonuçlarımız da bu kuralın bir istisnası değildir.
PMVP'deki iki taraflı hasar, damgalamaya dayalı tercih üretimini bozarsa, bu tür bir hasar, damgalamanın başlamasından sonra onun korunmasını tehlikeye atabilir mi? Aşağıdaki deneyler dizisi bu soruyu cevaplamak için kurulmuştur (McCabe ve diğerleri, 1982).
PMVG hasarı ve tercihin korunması
PMIG'deki hasarın izlerin korunması üzerindeki etkisi, şeması Şekil'de gösterilen bir deneyde incelenmiştir. 5.5. Tavuklar, önceki deneyde olduğu gibi aynı uyaranlar kullanılarak eğitildi , tek fark, doldurulmuş Banka tavuğunun sallanmaması, kutuyla aynı frekansta (1 dakikada 28 kez) döndürülmesiydi. Tavuklara, karanlıkta tutuldukları 70 dakikalık bir aralıkla ayrılmış 60 dakikalık iki periyot için dönen bir doldurulmuş tavuk veya kırmızı bir kutu verildi. Yaklaşık 3 saat sonra civcivlere anestezi uygulandı ve PMVG'nin iki taraflı imhası yapıldı ve sahte ameliyat edilen tavuklarda kafa derisi, beyine zarar vermiş kuşlarda olduğu gibi kesildi ve dikildi. Ek olarak, iki tavuk grubu daha tanıtıldı. Önceki deneyde (Bölüm 5.1), beyin hasarının spesifik olmayan sonuçları üzerinde kontrol yoktu. Bu nedenle Wulst hasarı olan bir grup tavuk gerekli kontrol olarak alındı. Bu, iki düşünce temelinde yapıldı . İki radyootografik çalışmanın hiçbiri (Horn ve diğerleri, 1979; Kohsaka ve diğerleri, 1979) bu alanda damgalama ile ilgili değişiklikler göstermedi ve Salzen ve diğerleri (Salzen ve diğerleri, 1975, 1978) bu alanda hasar olduğunu gösterdi . Evcil tavuklarda görsel Wulst'un damgalama üzerinde neredeyse hiçbir etkisi yoktur . Son olarak, Salzen ve arkadaşlarının ( 1975, 1978, 1979) lateral ön beyin hasar gördüğünde damgalamanın bozulduğuna dair verilerini test etmek için başka bir tavuk grubu tanıtıldı. Bu sonuç, damgalama sırasında önbeynin bu kısmına ilişkin radyootografik verileri sunan Kozaka ve diğerleri (Kohsaka ve diğerleri, 1979) tarafından da doğrulanmıştır . Bu nedenle, bir grup tavukta ön beynin yanal bölgesi hasar görmüştür. Ameliyattan sonraki gün , önceki deneyde kullanılan aynı test kullanılarak tüm civcivlerde tercih seviyeleri ölçüldü ve tercih puanları hesaplandı (s. 104).
tüm civcivler bozulmadan kaldığından, dört grubun tümü için yaklaşık puanların istatistiksel olarak aynı olması şaşırtıcı değildir (Tablo 5.2). Yaklaşma yanıtı, eğitim periyodu 2'de periyot 1'e kıyasla daha güçlüydü.
Tablo 5.2. Eğitim sırasında yakınlık puanları (McCabe ve diğerleri, 1982)
Tüm deney grupları arasında (beyin hasarı olan) önemli bir fark yoktur. Tüm gruplarda eğitimin 2. periyodundaki yaklaşım 1. periyoddakinden önemli ölçüde daha yüksektir (P<0.001). Kısaltmaların anlamı için, şek. 5.6.
Tercih göstergeleri, eğitim sırasında tavuklara sunulan nesneye bağlı değildi. Bu nedenle, kırmızı kutu ile eğitilen civcivler ve doldurulmuş tavuk için puanlar bir araya toplanmıştır. Dört grubun tümü için ortalama tercih puanları, Şekil 1 ve 2'de gösterilmektedir. 5.6. Beyni hasar görmüş üç gruptan yalnızca PMVH'si tahrip olmuş civcivler, sahte ameliyat edilen kontrol grubuna kıyasla daha düşük tercih puanlarına sahipti. Tercih puanları da hasarlı Wulst grubundan daha düşüktü. Diğer tüm Civciv gruplarının tercih göstergeleri rastgele olanlardan önemli ölçüde daha yüksekse
100

Resim: 5.6. Tavuklardan oluşan dört deney grubu için ortalama tercih puanları. LO - sahte olarak çalıştırılan; PMVG - PMVG'nin yok edilmesi; W , Wulst'un yok edilmesidir; LZM - beynin yanal bölgesinin imhası. Ortalama tercih puanları 50'yi aştı: sahte ameliyat grubunda 36.5±3.28 ( /=11.13 ; P<0.001); W grubu vardır 34.0±7.9 (/=4.30; P<0.01); LZM grubunda — 27,8±5,61 (/=4,96; Р<0,01). PMVH'si tahrip olan grupta tercih indeksi , 50'den (ortalama fark 14.4±7.53; /=1.91) önemli ölçüde farklı değildi ve sahte ameliyat edilen tavuklardan (/=3.15; P< 0.01) ve hasar görmüş tavuklardan anlamlı derecede düşüktü. Wulst (/=2.26; P<0.05). (McCabe ve diğerleri, 1982.)
çay seviyesinde, PMVP'si zarar görmüş civcivlerdeki puanlar 50'den önemli ölçüde farklı değildi. Diğer bir deyişle, PMVP'si zarar görmüş piliçler diğer gruplardan keskin bir şekilde farklıydı.
önemli ölçüde daha büyük bir tercih indeksi , gelişmiş tercihin korunduğu anlamına geliyorsa, PMVP'si zarar görmüş tavuklarda böyle bir hafıza izi bulunmadı. 50'lik bir tercih puanı, elbette, amnezinin, yani civcivlerin eğitim sırasında kendilerine sunulan nesnenin hafızasını kaybetmiş olduğunun mutlak kanıtı değildir. Örneğin, nesneyi mükemmel bir şekilde hatırlayabilirler, ancak hasar izlerin yeniden üretilmesini engelleyebilir. Bozulmuş hatırlamaya bağlı amnezi, özneye bir tür hatırlatma veya uyaran sunularak genellikle rahatlatılabilir. Örneğin Lewis ve diğerleri ( 1968), elektrokonvülsif amnezili farelerde üreme kusurlarını tespit etmek için bir yöntem geliştirdi. Bu yöntemde tanıma testinden önce bir “hatırlatıcı” kullanılmaktadır . Kullandığımız tercih testi , civcivin daha önce yalnızca biri sunulan iki uyarana verdiği tepkiyi ölçtüğü için bir tür tanıma testidir . Tercih testinde civcivler tanıdık ve yeni uyaranlara toplam 8 dakika maruz bırakıldı . Etkilerin sırası tabloda verilmiştir. 5.3. Civcivlerin yarısı (Tablo 5.3, Grup I)
Tablo 5.3. Bir tercih testinde civcivlere iki uyaranın (tanıdık ve yeni) sunulma sırası
McCabe ve diğerleri ( 1982)'deki tüm tavukların yarısı (Grup I) tablonun üst satırında gösterilen sırayla, diğer yarısı (Grup II) alt satırda gösterilen sırayla test edildi. Her 4 dakikalık test için kısmi tercih puanları, IOOX Tanıdık bir nesneye yaklaşmak (Tanıdık bir nesneye yaklaşmak) + (Yeni bir nesneye yaklaşmak) formülü kullanılarak hesaplandı.
önce tanıdık bir nesne gördü ve diğer yarısı (Tablo 5.3, grup II) yeni bir nesne gördü. Kısa süreli maruz kalma , hasar görmüş PMSH'li civcivlerde tanımayı geri getirirse, testin ilk yarısındaki civcivlerin tercih puanlarının ikinci yarıdakilerden farklı olması gerekir. Bu olasılığı araştırırken, her civciv için iki kısmi tercih puanı hesaplandı . Testin ilk 4 dakikası için bir gösterge ve sonraki 4 dakika için ikincisi hesaplandı. Testte tanıdık bir uyarana ilk maruz kalma civcive özelliklerini hatırlattıysa, o zaman tercih indeksi R (Tablo 5.3) tercih indeksi A'dan daha yüksek olmalıdır. Tercih indeksi R, tanıdık bir uyarıcı sunulan tavuklar için hesaplanmıştır . testin ikinci 2 dakikalık periyodunda nesne. ; tercih puanı A, testten önce karanlık bir inkübatörde tutulan ve bu nedenle hatırlatma testinden yararlanamayan civcivler içindir. PMVP'si zarar görmüş civcivlerde A ve G tercihlerinde anlamlı bir fark yoktu. Bu kriterler, zarar görmüş PMCH'li civcivlerde veya başka herhangi bir civciv grubunda hatırlama bozukluğuna dair kanıt sağlamaz .
tercih puanlarının düşmesinin başka nedenleri de vardır ; bunlardan biri test sırasında uyuşukluk olabilir. Uyuyorlarsa, o zaman elbette diğer beyin hasarlı kuşlara göre daha az hareketliydiler. Ancak, genel etkinlik
sırasındaki yaklaşım ve mesafe göstergelerinin toplamı olarak ifade edilen, tüm deney gruplarında anlamlı farklılık göstermemiştir.
Civcivlerin genel davranışlarını değerlendirmek için birkaç ek test kullanıldı. Tercih testinin ardından civcivler tekrar karanlık inkübatöre alındı. Her civciv daha sonra 25 cm yüksekliğinde karton bir silindire yerleştirildi, böylece civcivler duvarın ötesini göremedi. Silindirin zemininin üzerindeki duvardaki bir yarıktan çubuğa küçük kırmızı bir boncuk yerleştirildi (Şekil 5.7, H). Boncuk yavaşça sallandı ve boncuk gagalamaya kadar geçen süre kaydedildi. Ortalama göstergeler, tüm deney gruplarında önemli ölçüde farklılık göstermedi (Şekil 5.7, B). Bu sonuç önermektedir. beyin hasarı olan kuş gruplarının hiçbirinde görsel-motor koordinasyonunda ciddi ihlaller yoktu.
Bu sonucun daha fazla teyidi, gagalama tepkisi ile başka bir testte elde edildi. Sallanan boncuk testinden birkaç saat sonra, her bir civciv yere dağılmış darı serpilmiş ışıklı bir kuluçka makinesine yerleştirildi. Civcivler 30 dakika kuvözde bekletildi. Gagalama sayısı sayıldı ve civcivin kaç kez ıskaladığı not edildi. Gagalama sırasında isabetli isabetlerin yüzdesi hesaplandı . Tüm civcivlerde doğru gagalama sayısı yaklaşık %80'di; Bu parametrede gruplar arasında anlamlı fark yoktu .
Bu deneydeki tüm prosedürler tamamlandıktan sonra (ayrıca bkz. bölüm 6.1), tavuklar kurban edildi ve beyinleri çıkarıldı. Her gruptan rastgele dört beyin seçildi ve histolojik analizleri yapıldı. Seri kesitler yapıldı ve hasar hacmi, tavuk beyni atlasından alınan kesitlerin görüntülerine uygulanarak onarıldı. Hasar alanları bir planimetre kullanılarak ölçüldü ve hasarlı beyin dokusunun hacmi hesaplandı. Yukarıda açıklanan çalışmalarda olduğu gibi , SMVG'nin önemli bir kısmı yok edildi ve hasar bölgesi açıkça sınırlandırıldı (Şekil 5.8, L, bölümler IV ve V). Wulst'taki hasar (Şekil 5.8, B), esas olarak aksesuar hiperstriatumun rostral kısmını ve bunun yanı sıra kısmen altta yatan orta ve dorsal hiperstriatumu içermiştir . Ventral hiperstriatum bu yaralanmada neredeyse sağlam kaldı. Yanal hasar (Şekil 5.8, C) , özellikle serebral hemisferlerin orta kısmında olmak üzere ventral hiperstriatum ve neostriatumun yan bölgelerini ele geçirdi.
Baskı sürecindeki farklı hasarların sonuçları, hasarın büyüklüğündeki farklılıklarla, yani “kitlesel eylem” etkisi ile açıklanabilir mi? Ön beyin hacminin bir yüzdesi olarak ifade edilen hasar hacimleri , Şekiller 1 ve 2'de gösterilmektedir. 5.9. Bu üç ortalama neredeyse aynıdır, bu nedenle kütle etkisi açıklaması atılabilir.
PMGB'nin tek taraflı yıkımının bellek izlerinin oluşumu ve depolanması üzerindeki etkisi
beyin hasarına ilişkin çalışmaların daha fazla detaylandırılması sağlandı (Hot ve diğerleri, 1983). Önceki deneyler , SMVP'nin iki taraflı yıkımının, tanıdık bir nesne için tercih izlerinin oluşumunu ve korunmasını bozduğunu göstermiştir. Aşağıdaki deneyde, tercih edilen bir model geliştirmenin mümkün olup olmadığını bulmaya çalıştık.
140

.5MM
Resim: 5.8. Beyin dilim diyagramlarında gösterilen beyin hasarının histolojik resmi (1—VIII). Desenler ve: dilimleri yaklaşık 1 mm aralıklarla gösterin. En rostral dilim /'dir. Cheri, analiz edilen 4 kuşun her birinde yok edilen alanları gösteriyor. Gölgeli alanlar mk'yi belirtir; birden fazla beyinde yok edildi. A. PMVP'de hasar. B. Wulst hasarı. B. Beynin yanal bölgesinde hasar. 1 - ek hiperstriatum; 2 - ara hiperstriatum; 3 - dorsal hiperstriatum 4 - neostriatum; 5 - ventral hiperstriatum; 6 - ektotriyum; 7 - ventrikül; 8 - yükselen palaeotum; 9 - ilkel paleostriatum; 10 - arşiv. (McCabe ve diğerleri, 1982.)
Resim: 5.9. Ön beyin hacminin yüzdesi olarak ifade edilen hasarın hacmi . Sağ ve sol hemisferler için göstergeler birleştirilir. PMCH, Wulst (W) ve lateral beyin bölgesi (LZM) hasarlı gruplar için ortalama değerler verilmiştir . Veriler, her grubun dört üyesinin beyin ölçümlerine dayanmaktadır. (McCabe ve diğerleri, 1982.)
yalnızca bir PMVG'nin korunmasıyla. Bu koşullar altında bir tercihin geliştirilmesi mümkünse, bozulmamış SMVP'nin müteakip imhası sırasında üreme bozulur mu?
Kullandığımız araştırma yöntemleri, beyin bölgelerinin yok edildiği önceki deneylerdekiyle aynıydı. Bu deneylerdeki olayların sırası, Şek. 5.10. Karanlıkta bekletilen tavuklara anestezi uygulandı
B I—C=] rc n CZJ 1
Resim: 5.10. PMVP'de tek taraflı hasar olması durumunda öğrenme ve üreme üzerine deneydeki olayların sırası . İki tercih testi kullanıldı. A. Zarar görmüş PMV veya Wulst'lu civcivler. İlk başta hasar tek taraflı, sonra iki taraflıydı. B. Sham ameliyatlı kontrol grubu. (Horn ve diğerleri, 1983.) Yumurtadan çıktıktan yaklaşık 12 saat sonra çiftler halinde 8* . Bir çiftten bir tavuk, sahte olarak çalıştırılan bir kontrol görevi görürken, ikinci tavuk beynin dört bölümünden birine - sağ veya sol PMVG ve sağ veya sol Wulst - zarar verdi. Ameliyattan sonra civcivler tekrar karanlık inkübatöre alındı. Ertesi gün, bir çıkrığa yerleştirildiler ve her biri 1 saatlik iki periyot boyunca dönen kırmızı bir kutu veya doldurulmuş banka tavuğu gibi bir uyarana maruz bırakıldılar. Daha sonra civcivlere seri tercih testi uygulandı. Tamamlanmasından bir süre sonra, her bir tavuğa yeniden anestezi uygulandı ve kalan PMVG veya Wulst'a zarar verildi. Ertesi gün civcivler yaklaşık -88 saatlik olduklarında tercih tekrar test edildi.
Beyninin sol tarafında ilk hasar olan tavukların davranışı, Wulst ve PMVG lezyonları için eşit derecede doğru olan, sağ tarafında ilk hasar olan tavukların davranışlarından önemli ölçüde farklı değildi. Bu nedenle, beynin farklı yarım kürelerindeki bölümlerin her birine verilen hasarın sonuçları birleştirildi; aynısı kırmızı kutu veya doldurulmuş tavukla eğitilmiş tavuk grubu için de geçerliydi.
Tek taraflı PMSH veya Wulst yıkımından sonra eğitilen tavuklarda , tercih puanları rastgele seviyeden önemli ölçüde yüksekti ve kontrol grubunun sonuçlarından farklı değildi (Şekil 5.11, K). Tek taraflı yıkım durumunda öğrenmenin bozulmadığı açıktır. Deney grupları için yüksek tercih puanları , eğitim sırasında bunlara dahil olan civcivlerin daha az aktif olmalarına ve sahte olarak çalıştırılan kuşlara göre tanıdık uyaranlara çok daha az yaklaşmalarına rağmen elde edilmiştir (Horn ve diğerleri, 1983).
Oynatma için korunmuş bir PMVP'ye ihtiyaç duyulursa, hasarına tercih endeksinde rastgele bir seviyeye düşüş eşlik etmelidir. Elde edilen sonuç tam olarak budur. İkinci bir PMCH yaralanması olan civcivler için ortalama tercih puanı (Şekil 5.11, B, PMCH) 60.31 idi. Bu gösterge, 50'den bir standart hatadan daha az farklıydı ve bu nedenle rastgele seviyeden önemli ölçüde farklı değildi. Buna karşılık, Wulst'u ikinci kez hasar görmüş civcivler için ortalama tercih puanı , rastgele seviyeden önemli ölçüde daha yüksekti ve sahte ameliyat edilen civcivlerden neredeyse farklı değildi (Şekil 5.11b). Bu iki gruptaki indeksler, hasarlı PMVP'li gruba göre anlamlı derecede yüksekti .
edilen kısmi tercih ölçümleri (Tablo 5.3) , uyaranın tanıdık olup olmamasına bağlı değildi.

Resim: 5.11. Üç deneysel tavuk grubu için ortalama tercih puanları. Her ortalamanın sağında, kesikli çizginin üzerinde, bu ortalama 50'lik bir tercih etmeme puanıyla karşılaştırıldığında kritik fark (P=0,05) gösterilir. Ortalama, 50'den belirtilen kritik değerden daha fazla farklıysa, o zaman 50'den farklıdır. Bu testte elde edilen ortalama değerler arasındaki karşılaştırma için kritik farklar (P=0,05) da gösterilmiştir. LO — sahte ameliyat edilen tavuklar; W - Wulst hasarı; PMVG - PMVG'ye verilen hasar. Deney sırasında bazı civcivler öldü , bu nedenle orijinal sayıları (grup başına 48 kontrol ve 24 deney civciv) azaldı. (Horn ve diğerleri, 1983.)
yeni - önce sunulur. Veriler, PMCH'ye verilen hasarın etkisinin hatırlama bozukluğu ile sınırlı olmadığını bir kez daha gösterdi .
Bu çalışmaya ve yukarıda açıklanan iki beyin hasarı serisine dayanarak, imprinting yoluyla tercihi öğrenmek için en az bir PMVG'nin korunmasının gerekli olduğu ve hangi yarım kürede depolandığının önemli olmadığı sonucuna varılmıştır. Ayrıca, öğrenme yalnızca bir PMVP varlığında gerçekleşirse, üreme sırasında korunması gerektiği sonucuna varılmıştır.
Beyin hasarını kullanan diğer çalışmalar
ve radyootografik çalışmalardan ortaya çıkan varsayımları test etmek için yapılmıştır. Diğer yazarlar ayrıca ön beyin hasarının yaklaşımın çeşitli yönleri ve damgalama yanıtı üzerindeki etkisini de incelediler. Sonuçları birbiriyle uyumlu mu ?
Collias ( 1980) , evcil tavuklarda kısmi veya tam decerebrasyonun "erken sosyalleşme" üzerindeki etkisini inceledi . Operasyon, yumurtadan çıktıktan hemen sonra gerçekleştirildi ve ön beynin her iki yarım küresi de çıkarıldı. Kısmen decerebre edilmiş tavuklarda, ön beynin yaklaşık %75'i çıkarıldı; geri kalanı yarımkürenin bazal yapılarını ve paleostriatal çekirdek kompleksini içeriyordu (bkz. Şekil 5.8, 8 ve 9). Tavuk reaksiyonları iki şekilde test edilmiştir. İlkinde civciv, her iki ucunda bir hoparlör bulunan yaklaşık 3,5 m uzunluğunda bir koridora yerleştirildi. Bunlardan biri aracılığıyla, bir tavuğun gıdaklamasının kaydı çalındı. Tavuk koridorun ortasına yerleştirildi ve gıdaklama kaynağına yaklaşması kaydedildi. Normal kontrol civcivleri, yumurtadan çıktıktan sonraki ilk iki günde numunelerin %80'inde buna yaklaştı. Beyinsiz civcivler yürüyebildikleri ve yüksek bir ses duyulduğunda uyanık bir duruş alabildikleri halde ona yaklaşmadılar. Tam olarak decerebre edilmemiş civcivler, gırtlak kaynağına neredeyse normal civcivler kadar sık yaklaştı. Ek olarak, kısmen decerebre kuşlar, deneyci onlardan uzaklaşırken onu takip etti. Bu tamamen decerebre edilmiş tavuklarda gözlenmedi.
Bu "erken sosyalleşme" testleri, civcivlerin tanıdık ve yeni uyaranlar arasında seçim yapması gerekmediğinden, damgalama testleri değildir. Kollias tarafından kullanılan testler, tam olarak decerebre edilmiş civcivlerin yaklaşma tepkisi olmadığını, kısmen decerebre edilmiş civcivlerde ise yaklaşma tepkisi olduğunu göstermektedir.
Beyin hasarının damgalama üzerindeki etkisine ilişkin daha doğrudan bir çalışma, Salzen ve diğerleri ( 1975, 1978, 1979) tarafından yapılmıştır. Bu çalışmalar, posterolateral ön beyindeki hasarın tavuklarda tattan kaçınma tepkisini geliştirme ve sürdürme yeteneğinin tamamen kaybına yol açtığını gösteren Benowitz'in ( 1972) çalışmalarının devamıydı. Salzen bunu keşfetti
Resim: 5.12. K. Selzen ve Williamson tarafından derlenen tavuk beyninin stereotaksik atlasından bir dizi ön kesit . B. 14 tavukta ön beyin lateral lezyonları . Gölgeli alan, bu grupta iki taraflı olarak çıkarılan toplam doku hacmini gösterir ve siyah alan, vakaların %75'inde hasar gören alanı gösterir. Her dilimin altındaki rakamlar , belirtilen düzeyde hasarlı civciv sayısını gösterir. Stereotaksik koordinatlar (A) milimetre cinsindendir; A=9.6 en ileri koordinattır . 1 - dorsal hiperstriatum; 2 - ara hiperstriatum; 3 - ek hiperstriatum; 4 - ventral hiperstriatum; 5 - koku alma payı; 6 - neostriatum; 7 - ektotriyum; 8 - yükseltilmiş paleostriatum; 9 - ilkel paleostriatum; 10 - hipokamp; 11 - parahipokampal bölge; 12 - arşiv. (Salzen ve diğerleri, 1978.)
yarımkürenin yan kısmındaki hasar (Şekil 5.12) , tanıdık nesne için tercihin hem üretimini hem de kalıcılığını ortadan kaldırdı. Buna karşılık, görsel Wulst'u yok eden yarımkürenin rostral kısmındaki hasar , öğrenme sürecinin her iki yönü üzerinde çok az etkiye sahipti . Bu araştırmacı grubunun tüm deneylerinde PMVP etkilenmedi.
Bu nedenle, Wulst'un yok edilmesinin damgalama üzerinde çok az etkisi olduğu veya hiç etkisi olmadığı konusunda genel bir fikir birliği vardır. Salzen ve arkadaşlarının ( 1978) lateral ön beyin hasarının hafıza üzerindeki etkisi hakkındaki verileri bizim verilerimizle (bölüm 5.2) çelişiyor mu? Muhtemelen değil. Bu görüşün birkaç nedeni var: eğitimden sonra yan beyinleri hasar görmüş tavukların tanıdık bir nesneyi yenisine tercih ettiğini bulduk (Şekil 5.6). Bu davranış, seçim tepkisinin şans düzeyinde olduğu hasarlı PMVG'li civcivlerden farklıydı. Bununla birlikte, beynin yanal bölgesinde hasar olan tavuklarda, tercih göstergeleri , hem sahte ameliyat edilen grubun hem de PMVP'nin hasar gördüğü grubunkinden önemli ölçüde farklı değildi , yani yanal hasarlı grup, arada kalan ortalama değerler verdi. diğer iki grubun özelliği olan uç değerler. Bizim çalışmamızda lateral beyindeki hasar, Salzen ve arkadaşlarının ( 1978) çalışmasına göre biraz daha az yaygındı . Onların deneylerinde hasar, bizim deneylerimizde etkilenmeyen bir yapı olan arşistriatumun bir bölümünü kapsıyordu (McCabe ve diğerleri, 1982). Lezyonların boyutu ve lokalizasyonundaki farklılıklar , Şekiller karşılaştırıldığında kolayca görülebilir. 5.8, B ve 5.12, B. Böylece, Şek. 5.8, B* archistriatum'u ele geçirdi, ardından bu gruptaki ortalama tercih, PMCH'si zarar görmüş grubun karakteristik değerlerine daha yakındı. Tabii ki, bu sorun deneysel olarak çözülebilir , ancak PMCH'nin beynin ve arkistriatumun yanal bölgesine yansıtıldığını not etmek önemlidir (Bradley ve Hom, 1979; Bradley ve diğerleri, 1985).
İki çalışma grubu arasında başka farklılıklar da vardır. Salzen ve meslektaşları ameliyat edilen her tavuğu tavandan sarkıtılmış bir nesne içeren bir kutuda yetiştirdiler. Sarı bir sünger veya yeşil kumaştan bir top olan nesne , kutunun ortasında serbestçe sallanıyordu. Civcivler 3. günden 10. güne kadar bu kutularda tutuldu. Bu nedenle, bu çalışmadaki civcivler, Bölüm 1'de açıklanan deneylerdeki civcivlerden daha yaşlıydı. 5.2 ve 5.3 ve çok daha uzun süre tamamen farklı uyaranlar sunuldu. Bu faktörler, damgalama sürecinin nöronal temelinin analizinde büyük önem taşır (Bölüm 7 ve 8).
Takamatsu ve Tsukada (Takamatsu ve Tsukada, 1985), ön beyinleri hasar görmüş ördek yavrularını kullanarak radyootografik çalışmalarına (Kohsaka ve diğerleri, 1985) devam ettiler. Görünüşe göre PMSH'ye karşılık gelen ventral hiperstriatumun orta kısmını içeren yaralanmalar, sahte çalıştırılan ördek yavrularında gözlenmeyen aşağıdaki reaksiyonun ihlaline yol açtı.
Beyin İmha Değerlendirmesi
Beynin, bir kaza sonucu veya deneysel bir amaçla patolojik bir süreçle hasar gördükten sonra kalan işlevlerini inceleme yöntemi, bir sinirbilimcinin cephaneliğindeki en kaba yöntemlerden biridir. Bu yöntemin bazı sınırlamalarından daha önce bahsedilmişti; bu sınırlamalar, Gregory ( 1961 ) tarafından yapılan bir incelemede de kapsamlı bir şekilde tartışılmıştır . Bunu not ettikten sonra, eğer bu yöntem kullanılmasaydı sinirbilim tarihinin çok farklı olabileceğini ve beyin hakkındaki bilgilerimizin daha sınırlı olabileceğini de kabul etmek gerekir. Bazı durumlarda bu mümkün olan tek yöntemdir ve onun yardımıyla yapılan gözlemler genellikle sonraki, daha incelikli analizler için temel teşkil eder. Bunun örnekleri çoktur. Omurilik fonksiyonlarının deneysel analizine yönelik ilk yaklaşımlar esas olarak bu yönteme dayanıyordu (Liddell, 1960). İnsanlarda asimetrik beyin işleviyle ilgili ilk çalışmalar , tek taraflı kortikal lezyonların konuşma veya sözlü ve yazılı dili anlamadaki bozukluklarla olan korelasyonlarına dayanıyordu (Broca, 1865; Wernicke, 1874; Dejerine, 1891). Daha yakın zamanlarda, Scoville ve Milner'ın ( Scoville ve Miller, 1957) hasarlı beyin çalışmaları , hipokampusa derin bir ilgi uyandırdı ve bu oluşumun işlevlerine ilişkin çok sayıda verimli deneysel çalışmayı teşvik etti (Ciba Foundation Symposium, 1978; Seifert, 1983). Primat beyninin alt temporal lobunun görme odaklı davranıştaki rolü ilk olarak Chow (Chow, 1952) ve Mishkin ve Pribram'ın (Mishkin, 1954; Mishkin ve Pribram, 1954) deneylerinde bu lobun çıkarılmasıyla açıkça gösterilmiştir. Bu yazarlar tarafından elde edilen veriler , çok sayıda elektrofizyolojik çalışmanın temelini oluşturdu (örneğin, bkz . Bruce ve diğerleri, 1981; Perrett ve diğerleri, 1982). Skotom alanında artık görmenin keşfi, beyin hasarının artık görsel işlev üzerindeki etkisinin derinlemesine bir analizine (Weiskrantz, 1980; Zihl ve Werth, 1984) ve hasarın etkilerinin gözlemlenmesine dayanıyordu. insanlarda parietal lob (Critchley, 1953) incelikli elektrofizyolojik maymun dikkat çalışmalarının temelini attı (Lynch ve diğerleri, 1977; Wurtz ve diğerleri, 1980; Bushnell ve diğerleri, 1981).
İşlev bilgisi yalnızca beyin hasarlı deneylerden elde edilen bilgilere dayansaydı , o zaman olası yorumlar elbette çok sınırlı olurdu. Damgalama çalışmasında, bu bölümde ayrıntılı olarak tartışılan beyin hasarı deneylerinin sonuçları, PMVP'nin bu tür davranışlardaki işlevi hakkındaki tek bilgi kaynağı değildi . Bu sonuçlar , PMVP'nin damgalama sürecinde kritik bir rol oynadığına dair, esas olarak biyokimyasal verilere dayalı olarak zaten yapılmış olan varsayımları doğruladı . Bu süreçteki ciddi bozukluklar, el-göz koordinasyonu , lokomotor aktivite, uyanıklık seviyesi veya görsel farklılaşma yeteneğindeki kusurlara bağlı değildir, çünkü PMVG'siz tavuklar, sahte çalıştırılan kuşlar kadar algı ve motor aktivite testlerinde performans göstermiştir (bkz. ayrıca Bölüm 6-8). Hasarlı bir PMVG'ye sahip civcivlerin hatırlama bozukluğundan mustarip oldukları neredeyse hiç varsayılamaz (s. 111). Gentle ve diğerleri ( 1978), yetişkin evcil tavuklarda ventral hiperstriatum içeren lezyonların, korkutucu bir uyarana tepkilerini azalttığını varsaydılar. Belirli bir görsel nesneye damgalanmış civcivler diğer nesnelere yaklaşmazlar ve hatta onlardan aktif olarak kaçınabilirler. PMVG'ye verilen hasar civcivlerin korku duygusunu azaltırsa, o zaman bir tercih testinde yeni bir nesneye tepkide bir artış beklenebilir; ancak durum böyle değildi. PMVG'nin yaralanması, civcivlerin yeni bir nesneye yaklaşma tepkisini etkilemedi (Tablo 5.1; ayrıca bkz. Horn ve McCabe, 1984). Bu nedenle, hasarın sonuçlarının korkuda genel bir azalma ile açıklanması olası değildir. Başka bir açıklama da, etkilenen civcivlerin uygun şekilde motive olmadıkları için tanıdık nesneyle ilgilenmemeleridir. Bu argümanı çürütmek zordur, ancak bazı veriler davranışlarının bu açıklamasının doğru olmadığını göstermektedir. Dolaylı kanıtlardan biri, Bölüm 1'de açıklanan "iki günlük" biyokimyasal deneylerden gelmektedir. 3.2. Bu deneylerde, dört grup tavuk, yaşamın ilk gününde farklı süreler için eğitildi. İkinci gün tüm gruplara 60 dakika antrenman yaptırıldı. 2. günde rostral ön beyin çatısına radyoaktif urasil eklenmesinin, 1. günde en uzun süre antrenman yapan civcivlerde en düşük olduğu bulundu. Düşük katılım düzeyi motivasyondaki bir düşüşü yansıtabilir ve bu da tavuğun davranışının bazı yönlerinde kendini göstermelidir. 1. günde en uzun süre eğitilen civcivler, uyaran hakkında en fazla bilgiyi almış olmalıdır (Şekil 3.5,2>). Ertesi gün onu gördüklerinde ona "ilgilerini kaybedebilirler" ve bu nedenle ona daha az aktif bir şekilde yaklaşabilirler. Ancak bu olmadı. Aslında (Şekil 3.5, B), önceki gün daha uzun süre eğitilen kuşlar, diğer civciv gruplarına göre tanıdık bir nesneye doğru daha uzun mesafe koşma eğilimindeydiler, ancak bu eğilim istatistiksel olarak anlamlı değildi. PMCH'si zarar görmüş civcivlerin motivasyon bozukluğu yaşamadığının bir başka göstergesi de normal gagalama tepkilerini ve beslenme davranışlarını sürdürmeleridir. Diğer bazı davranışlar da bu hasardan etkilenmez (bkz. Bölüm 6 ve Kısım 7.4, 8.1 ve 8.4.1).
PMVP'deki hasarın baskı üzerindeki etkisi, iz depolamanın ihlali olarak yorumlanabilir; bu durumda öğrenme ve ardından gelen tanıma testleri sırasında bilgi beynin bu bölgesinden geçer ancak beyne kaydedilmez . Bu durumda radyootografik çalışmaların sonuçları (Hot ve diğerleri, 1979), başka bir yerde saklanan bilgi alındığında MVG'den geçerse "kullanımdaki hipertrofi "nin sonucu olabilir . Böyle bir bölgede, örneğin içinde nöron elementlerinin büyüme süreçleri meydana gelirse, artan biyokimyasal aktivite tespit edilebilir. Bununla birlikte, radyootografik çalışmalar , iki görsel projeksiyon alanı ve motor aktiviteyi düzenleyen alanlar dahil olmak üzere beynin diğer birçok bölümünü analiz etmiştir. Bu alanların hiçbirinde, PMSH'ye özgü bir resim gözlenmedi. Sonuç olarak, nöronların kendi içinde "kullanımı", yerel radyootografik değişikliklere yol açmaz. Bu düşünceler, bu hipotezin olasılığını azaltır.
Damgalamayla ilgili ilk biyokimyasal çalışmanın (Bölüm 2) sonuçlarını yorumlamak kesinlikle çok zordu. Beynin işleyişine ilişkin bir dizi başka biyokimyasal göstergeyi ortaya çıkaran bir dizi kontrol deneyinin yanı sıra beyin hasarının yardımıyla, başlangıçtaki belirsizlikleri kademeli olarak ortadan kaldırmak mümkün oldu. Genel olarak, elde edilen veriler, PMIG'nin baskı sırasında tercihlerin oluşması için gerekli olan bilgilerin depolanması için kritik öneme sahip olduğunu düşünmemizi sağlar . Bu veriler ayrıca, civcivlerde görsel bir nesnenin etkisi altında, nesne ezberlemenin bazı yönlerinin temeli olan SMVG'nin nöronal organizasyonunda değişiklikler olduğunu göstermektedir . PMVP'nin bilgi depolamaya katılımının yalnızca damgalama için mi yoksa diğer öğrenme biçimleri için mi gerekli olduğu sorusu bir sonraki bölümde ele alınacaktır. Baskılama açısından, PMVP'deki sözde depolama işlevinin, civciv baskısı sırasında yaklaşma tepkisinin yönlendirildiği belirli bir uyaran türüyle sınırlı olup olmadığı da açık bir sorudur (bkz. Bölüm 8).
Özet
Bölümde açıklanan deneylere dayanarak. Şekil 2-4'te, ön beynin belirli bir bölümünün, yani PMCG'nin, civciv kendisine görsel bir nesne sunulduğunda aldığı bilgileri depolamak için kritik olduğu öne sürüldü. Bu varsayımdan, PMVP yok edildiğinde, baskı sırasında tercihlerin geliştirilmesi ve saklanmasının imkansız olması gerektiği sonucu çıktı. Bu bölümde açıklanan deneyler, PMVP'nin çift taraflı tahribatı durumunda civcivlerin eğitimden önce tanıdık bir nesne için bir tercih geliştirmediğini göstermektedir. Bu bölgenin iki taraflı tahribatı eğitim tamamlandıktan sonra yapılırsa, tercihin korunması büyük ölçüde ihlal edilir. Çeşitli olasılıkların elenmesi yöntemiyle, PMCH'nin anneyi tercih etme ve takip etme gelişimi ile doğrudan ilgili bilgileri depolamak için kritik olduğu sonucuna varılabilir .
EDEBİYAT
Benowitz L. Ön beyin ablasyonlarının civcivlerde kaçınma öğrenme üzerindeki etkileri, Physiol. Behav., 1972, 9, 601-608.
Bradley P., Horn G. Civcivlerde hyperstriatum ventrale'nin götürücü bağlantıları, J. Anat., 1979, 128, 414-415.
Bradley P., Davies DC, Horn G. Evcil civcivde (Gallus domesticus) hyerstriatum ventrale'nin bağlantıları , J. Anat., 1985, 140, 577-589.
Broca P. Sur la dil fakültesi makalesi, Bull. sos. Antropol., Paris, 1865, 6, 337-393.
Bruce C., Desimone R., Gross CG Makağın üst temporal sulkusundaki çok duyusal bir alandaki nöronların görsel özellikleri, J. Neurophysiol., 1981, 46, 369-384.
Bushnell MC, Goldberg ME, Robinson DL Maymun serebral korteksinde görsel tepkilerin davranışsal gelişimi. I. Posterior parietal kortekste seçici görsel dikkat ile ilgili modülasyon, J. Neurophysiol., 1981, 46, 755-772.
Chow KL Görsel olarak yönlendirilen davranışla ilgili olarak çağrışımsal korteksin seçici ablasyonunun diğer öğrencileri, J. comp. fizik. PsychoL, 1952, 45, 109-118.
Ciba Vakfı Sempozyumu. Septo-hipokampal sistemin işlevleri, Ciba Vakfı Sempozyumu, 1978, no. 58, Excerpta Medica, Amsterdam.
Collias NE Bazal telensefalon, civcivlerde erken sosyalleşme için yeterlidir. fizyol . Behav., 1980, 24, 93-97.
Critchley M. Parietal loblar. Arnold, Londra, 1953.
Gentle MJ, Wood-Gush DGM, Gordon J. Gallus domesticus'ta hiperstriatal ablasyonun davranışsal etkileri , Behav. Süreç, 1978, 3, 137-148.
Gregory R.L. Bir mühendislik problemi olarak beyin. İçinde : Hayvan davranışındaki güncel problemler (ed. EH Thorpe ve OL Zangwill), 1961, s. 307-330, Cambridge University Press, Cambridge.
Horn G., McCabe BJ Önyargılar ve tercihler. Baskı üzerindeki etkiler
civciv beynine lezyonlar, Anim. Behav., 1984, 32, 288-292.
Horn G., Rose SPR, Bateson PPG Merkezi sinir sisteminde harcama ve plastisite, Science, 1973, 181, 506-514.
Horn G., McCabe B. J. Bateson PPG Damgalamadan sonra civciv beyninin otoradyografik bir çalışması, Brain Res., 1979, 168, 361-373.
Horn G., McCabe BJ, Cipolla-Neto J. evcil civcivde damgalama: hiperstriatum ventrale'nin her iki tarafının edinme ve tutmadaki rolü, Exp. Brain Res., 1983, 53, 91-98.
Kohsaka S.-L, Takamatsu K., Aoki E., Tsukada W. Civciv beyninin [14C] 2-deoksiglukoz kullanılarak imprinting sonrası metabolik haritalaması , Brain Res., 1979, 172, 539-544.
Lewis D. L, Misanin JR, Miller RR Amnezi sonrası hafızanın kurtarılması. Nature, Londra., 1968, 220, 704-705.
Liddell EGT Reflekslerin keşfi. Clarendon Press, Oxford, 1960.
Lynch JC, Mountcastle VB, Talbot WH, Yin TCT Yönlendirilmiş görsel dikkat için parietal lob mekanizmaları, J. Neurophysiol., 1977, 40, 362-389.
McCabe B. L> Horn G., Bateson PPG Civciv ön beyninin kısıtlı lezyonlarının damgalama sırasında evlat tercihlerinin edinilmesi üzerindeki etkileri, Brain Res., 1981, 205, 29-37.
McCabe BJ, Cipolla-Neto J „ Horn G., Bateson PPG Baskıdan sonra civcivin hiperstriatum ventralinde yer alan bilateral lezyonların amnezik etkileri , Exp. Brain Res., 1982, 48, 13-21.
Mishkin M. Temporal lobun kısmi ablasyonunu takiben görsel ayrım performansı. II. Ventral yüzey vs. hipokampus, J. komp. fizik. Psychol., 1954, 47, 187-193.
Mishkin M., Pribram K. H. Temporal lobun kısmi ablasyonunu takiben görsel ayrım performansı. I. Ventral vs. yanal, J. komp. fizik. Psychol., 1954, 47, 14-20.
Perrett DI, Rolls ET, Caan W. Maymun temporal korteksindeki yüzlere yanıt veren görsel nöronlar, Exp. Brain Res., 1982, 47, 229-238.
Salzen EA, Sluckin W. Evcil kümes hayvanlarında aşağıdaki yanıtın insidansı ve yanıt verme süresi, Anim. Behav., 1959, 7, 172-179.
Salzen EA, Parker DM, Williamson AJ Evcil civcivlerde baskıyı önleyen bir ön beyin lezyonu, Exp. Brain Res., 1975, 24, 145-157.
Salzen EA, Parker DM, Williamson AJ Evcil civcivlerde ön beyin lezyonları ve imprinting retansiyonu, Exp. Brain Res., 1978, 31, 107-116.
Salzen EA, Williamson A. Parker DM Evcil civcivlerde ön beyin lezyonlarının doğuştan gelen ve damgalanmış renk, parlaklık ve şekil tercihleri üzerindeki etkileri, Behav. İşlem., 1979, 4, 295-313.
Scoville WB, Milner B. Bilateral hipokampal lezyonlardan sonra son hafıza kaybı, J. Neurol. beyin cerrahisi. Psikiyat., 1957, 20, 11-21.
Seifert W. (ed.) Hipokampusun Nörobiyolojisi, Academic Press, Londra, 1983.
Takamatsu K., Tsukada Y. Civciv ve ördek yavrusu damgalamanın nörobiyolojik temeli. In: Nörobilimlerde Perspektifler: molekülden akla. (ed. Y. Tsukada), 1985, s. 187-206, Springer, W. Berlin.
Weiskrantz L. Kalan deneyim çeşitleri. litre. J. exp. Psychol., 1980, 32, 365-386.
Wernicke C. Der aphasische Symtomen Complex. Cohn ve Weigert, Breslau, 1874.
Wurtz B. H., Goldberg ME, Robinson DL Maymundaki görsel tepkilerin davranışsal modülasyonu: dikkat ve hareket için uyaran seçimi. Program Psychobiol. fizik. Psychol., 1980, 9, 43-83.
Zihl J., Werth R. “Görme” çalışmasına katkı. I. Postgenikulat alan defektleri olan hastalarda sakkadik lokalizasyon için başıboş ışık açıklanabilir mi? Nöropsikoloji, 1984, 22, 1-11.
Zeuner FE Evcilleştirilmiş hayvanların tarihi. Hutchinson, Londra, 1963.
BÖLÜM 6
BASKI VE İLİŞKİLİ ÖĞRENME
PMVG'si zarar görmüş civcivler, daha önce gördükleri bir nesnenin hiçbir özelliğini hatırlamamaları dışında, oldukça normal davranırlar. Bununla birlikte, PMVP'deki hasarın, incelediğimiz basit el-göz koordinasyonu gözlemleriyle tespit edilemeyen ince davranış bozukluklarına neden olması mümkündür . Örneğin, beyin hasarının tavukların küçük parlak nesneleri veya yiyecek parçalarını gagalama yeteneğini bozmadığı, ancak çevrelerindeki dünya hakkında hiçbir şey öğrenemedikleri kolayca düşünülebilir. Bu olasılığı araştırmanın bir yolu , PMCH'deki hasarın damgalamanın yanı sıra diğer öğrenme türlerini bozup bozmadığını bulmaktır. Bu amaçla, şu soruyu cevaplamaya çalıştıkları iki dizi deney gerçekleştirildi : PMSH'deki hasar ilişkisel öğrenmenin ihlaline yol açar mı?
Sıcak hava ile takviye edildiğinde reaksiyon gelişimi
Hayvanlarda belirli bir tepki oluşturmak için genellikle yiyecek veya su pekiştireç olarak kullanılır . Bu pekiştirici uyaranlar, üretilen tepkilerde bir artışa veya artışa yol açar . Bununla birlikte, yaşamın ilk birkaç gününde yumurta sarısını besin kaynağı olarak kullanan civcivlerin davranışları üzerinde yiyecek ve suyun çok az etkisi vardır. Zolman ( 1968, 1969), ısı takviyesinin civcivlerde yaklaşma tepkisini geliştirmek için başarılı bir şekilde kullanılabileceğini göstermiştir. Deneylerimizin ilkinde, 3 günlükten küçük civcivler kırmızı bir daireye yaklaşmaları için eğitildi. Plastik bir çubuğa monte edilen daire, eğitimin yapıldığı iki odadan birine belirli bir sıra ile yerleştirildi. Tüm deney düzeneği, sıcaklığın 10°C'de tutulduğu bir odaya yerleştirildi . Doğru yaklaşma reaksiyonu, bir kızıl ötesi lambadan 15 saniye boyunca sağlanan ısı ile desteklendi . Her tavuğa 20 örnek verildi. Civcivler öğrendikçe, doğru seçimi yapma olasılıkları önemli ölçüde arttı . Bu sonuç, uygun koşullar altında civcivlerin belirli bir uyarana yaklaşmayı ısı ile ilişkilendirdiğini göstermiştir. PMVG'si zarar görmüş civcivler benzer bir sorunu çözebilir mi?
Bu sorunun incelendiği tavuklar, Bölüm 1'de açıklanan deney dizilerinde de kullanılmıştır. 5.2 (McCabe ve diğerleri, 1982). Bu deneylerin özünü kısaca hatırlayalım. PMVG'nin iki taraflı yıkımı, bir tavuk grubunda, diğerinde Wulst ve üçüncüsünde beynin yanal bölgesinde gerçekleştirildi. Yaralanmadan önce, sahte ameliyat edilenler de dahil olmak üzere tüm tavuklara kırmızı bir kutu veya doldurulmuş tavuk verildi. Tanıdık nesne tercihi yalnızca zarar görmüş PMVP'li civcivlerde bozuldu. Bu grubun tavukları, çeşitli yaralanmalara sahip diğerleri gibi, hareket eden kırmızı boncuğu sahte ameliyat edilen tavuklar kadar hızlı ve doğru bir şekilde gagaladılar; kuluçka makinesinin zeminine dağılmış darı tanelerinin gagalama sayısı veya doğruluğu açısından gruplar arasında önemli bir fark yoktu.
Gagalama testinin sonunda civcivler 15 saat süreyle karanlık inkübatöre geri döndürüldü. Daha sonra civcivler 2,5-3 günlük olduklarında deneyin bir sonraki aşamasına geçildi . Tavuklar, sıcak hava ile takviye edilirken iki görsel görüntüyü ayırt edecek şekilde eğitildi. Deney, 12°C'de soğuk bir odada gerçekleştirildi. Kullanılan kurulum, Şek. 6.1. Sabitlemek için verildiği odaların her birinin üzerinde , içinden kutuya bir sıcak hava akımının yönlendirilebileceği bir saç kurutma makinesi vardı. Karanlık kutudaki tavuklar, inkübatörden deney odasına alındı ve oda sıcaklığında bir süre kutuda bırakıldı . Ardından eğitim başladı. Her kuşa 24 örnek verildi . Kuşların yarısı mavi-siyah görüntüye yaklaştıklarında diğer yarısı da sarı-siyah görüntüye yaklaştıklarında takviye edilmişlerdir. Güçlendirilmiş uyaranın konumu (24 denemenin her birinde sağ veya sol bölmede), aparatın belirli bir tarafına (sol veya sağ) koşullu tepkiler geliştirme olasılığını veya yanıtların mekansal değişimini önleyen bir şemaya göre belirlendi. .
Koridora girmeden önce her grubun başlama odasında kaldığı süre aynıydı (Tablo 6.1). Şek. 6.2, her bir tavuk grubu için , ilk ve son numunelerde doğru ve yanlış reaksiyon gösteren kanatlıların sayısını gösterir . İlk denemede, seçim tüm gruplarda rastgele seviyeden farklı değildi (Şekil 6.2, L). Buna karşılık , son denemedeki doğru reaksiyonların sayısı, tüm gruplarda rastgele seviyeden önemli ölçüde daha yüksekti (Şekil 6.2, B).
Resim: 6.1. Termal takviye üzerinde şartlandırılmış reflekslerin geliştirilmesi için aparat . Soldaki şemadaki boyutlar, belirtilen 76 cm'lik ölçeğe karşılık gelir, takviyelerin sağlandığı kutunun zemini ve yan duvarları beyaz, koridor ve başlangıç kutusu siyahtır. Görsel uyaranlar - 10 cm çapında açık renkli daireler - şeffaf arka duvarlara yansıtıldı. Bir daire , aynı genişlikte sarı şeritlerle ayrılan 1,3 cm genişliğinde koyu şeritlerden oluşuyordu ; ikincisi - mavi zemin üzerine 2,5 cm çapında siyah dairelerden. (McCabe ve diğerleri, 1982.)
Her gruptaki civcivlerin yaklaşık yarısı, arka arkaya sekiz doğru yanıttan oluşan öğrenme kriterine ulaştı. Bu kritere ulaşmadan önce gruplar arasındaki örneklem sayısı önemli ölçüde farklılık göstermedi (Tablo 6.1). Bu nedenle, çeşitli göstergelere bakılırsa, tüm tavuk grupları 24 deneme sırasında eş zamanlı görsel seçim becerisi geliştirdi; gruplar arasında anlamlı fark yoktu.
Bu sonuçlar, PMVP'ye verilen hasarın genellikle tavukların algısını veya motor tepkilerini bozmadığına dair diğer testler temelinde varılan sonucu tamamen doğruladı. Ancak bu deneylere dayanarak PMVP'nin termal güçlendirme sırasında görsel farklılaşmanın geliştirilmesinde yer almadığını söylemek mümkün müdür? Sonuçlar bununla çelişmese de başka yorumlar da mümkündür. Bu görevde civcivlerin eş zamanlı görsel ayrım yapmaları gerekir ki bu onlar için damgalamanın gerektirdiği sıralı ayrımdan daha kolay olabilir . Ek bir komplikasyon , termal takviye ile şartlandırılmış bir refleksin gelişiminin, damgalama prosedüründen 30 saat sonra gerçekleştirildiği
gerçeğinde yatmaktadır.
Tablo 6.1. Eşzamanlı görsel ayrımcılığın geliştirilmesi (McCabe ve diğerleri, 1982)
İkinci sütunda verilen kriter, sekiz ardışık doğru cevaptı; Bu kriteri sağlayan civcivlerin oranı parantez içinde gösterilmiştir. Ne kritere ulaşan civcivlerin oranı ne de kuluçka için gerekli numune sayısı gruplar arasında önemli ölçüde farklılık göstermedi. Fırlatma kutusundan çıkıp koridor boyunca hareket etmeye başlamak için gereken sürenin ortalaması 24 deneme üzerinden alındı. Civciv pekiştireç verilen kutuya 120 sn içinde girmezse 120 sn gösterge atanmıştır. Ortalama değerler verilmiştir. Gruplar arasında önemli bir fark yoktur. Grup kısaltmalarının açıklaması için, şek. 5.6.
tinga. Civcivler olgunlaştıkça, uyaranları daha etkili bir şekilde veya başka bir şekilde ayırt etmeyi öğrenebilirler. Kuşkusuz, bu zorlukların üstesinden gelmek için, PMCH'ye verilen hasarın, damgalama sırasında ve edimsel şartlandırılmış reflekslerin gelişimi sırasında bilginin sabitlenmesi üzerindeki etkilerinin daha doğrudan karşılaştırılmasına izin verecek ek bir deneye ihtiyaç vardı.
Resim: 6.2. Eşzamanlı görsel farklılaşmanın geliştirilmesi. 24 örneklik bir seride ilk (A) ve son (B) örneklerde doğru (soldaki çubuklar) ve yanlış (sağdaki çubuklar) yanıtlara sahip civcivlerin sayısı . Her bir sütun çiftinin altında, doğru ve yanlış reaksiyonların eşit olasılıklı sıfır hipotezinden farklılıkların önemi gösterilmektedir; ND - fark önemli değil. Veriler tüm gruplar için sunulmuştur (Şekil 5.6'nın başlığına bakın). (McCabe ve diğerleri, 1982.)
140
Bir nesneyi görme yeteneği ile pekiştirildiğinde tepkilerin gelişimi
ördek yavruları (Anas rubripes tristis ve Anas platyrhynchos) o nesneyi görmek için diski gagalamayı öğrenirler (Peterson, 1960). Bu çalışmalar, belirli bir nesneye ördek yavrusu damgası vurduktan sonra, bu nesnenin pekiştirme özelliği kazandığını göstermiştir (ayrıca bkz. Campbell ve Pickleman, 1961; Hoffman ve diğerleri, 1966). Baskı yapan bir uyarıcı olarak etkili olan bir nesnenin daha sonra etkili bir koşullu pekiştireç haline geldiği de gösterilmiştir (Bateson ve Reese, 1969). Bu araştırmacılar, bir günlük civcivlerin yanıp sönen kırmızı veya mavi ışıkları görmek için pedala basmayı çabucak öğrendiklerini keşfettiler. Bir reaksiyon geliştirme kriterine ulaşıldığında, tavuklara bir seçim testi verildi. Koridorun uçlarına takviye görevi gören kırmızı veya mavi titreşen bir ışık yerleştirildi. Tavuk koridorun ortasına yerleştirildi ve yaklaşma tepkileri kaydedildi. Tavukların eğitim sırasında gördükleri uyarana yaklaştıkları ortaya çıktı . Bu deneyler, görsel bir nesnenin, eğitimden önce görsel deneyimi olmayan tavuklarda davranışsal tepkilerin pekiştirilmesi olarak hizmet edebileceğini gösterdi. Ek olarak, deney , civcivlerin pedala basmak ile titreyen bir ışığın görüntüsü arasında çağrışımsal bir bağlantı geliştirdikçe, nesnenin özelliklerini hatırladıklarını ve daha sonra onu tanıyabildiklerini gösterdi. Her iki süreç - tanıma ve bağlanma - aynı anda meydana geldiğinden, bu öğrenme biçimi, bu süreçlerdeki ayrışmanın PMSH'nin yıkımından kaynaklanıp kaynaklanmadığını bulmak için kullanılabilir (Johnson ve Hoth, 1984, 1986).
Edimsel öğrenme için kurulum, şekil 2'de gösterilmiştir. 6.3. Kutunun üç tarafı siyah mat boya ile boyandı. Dördüncü duvar, önünde takviye edici bir nesne olan kırmızı bir kutu veya doldurulmuş bir tavuk olan tel örgüden yapılmıştır. Kutunun zemininde siyah ve beyaz kareli dizilmiş iki göze çarpan pedal vardı . Tavuklar pedallara basarak basabiliyordu. Pedallardan birine basılması bir röleyi kapatarak, civciv pedal üzerinde kaldığı sürece güçlendirici nesnenin yanmasına ve dönmesine neden oldu. "Açık" nesne tavuk için çekici hale geldi ve ona yaklaşmaya çalışarak pedaldan indi . Sonuç olarak, nesnenin aydınlatması ve hareketi durdu . Başka bir pedala basmanın hiçbir etkisi olmadı. Ağırlıklı olarak sağ veya sol pedala basma eğilimini ortadan kaldırmak için , "aktif" pedalın konumu

Resim: 6.3. Metal bir ağ duvarının önüne yerleştirilen güçlendirici uyaranlardan biri ile edimsel öğrenme için bir aparatın şeması . Bu duvarın boyutları 42x30 cm'dir.Uyarıcı aparatın önüne 35 cm uzaklıkta yerleştirilmiştir (Bateson ve Reese, 1969).
farklı kuşlar için sürekli değişiyordu. Herhangi bir kuş için aktif pedal her zaman solda veya sağdaydı. Bu nedenle, civcivlerin dönen bir nesneyi görerek güçlenmeleri için belirli bir yere gitmeleri gerekiyordu .
Deneyde iki grup tavuk kullanıldı - sahte olarak çalıştırılan ve iki taraflı PMVP yıkımı yapılan. Civcivler yaklaşık 12 saat olana kadar karanlıkta tutuldu.Sahte ameliyat edilen kuşlara anestezi uygulandı, kafa derisi kesildi ve dikildi. PMVG'nin yok edilmesi yukarıda açıklanan yöntemle gerçekleştirilmiştir (Bölüm 5.1). Operasyon sırasında civcivler önceki gibi kodlanmış ve sonraki tüm işlemler deneyci tavuğun hangi gruba ait olduğunu bilmeden gerçekleştirilmiştir . Edimsel reflekslerin gelişimi tavuklar ameliyattan tamamen kurtulduklarında ve 20-24 saat yaşına geldiklerinde başladı.Tavukların bir yarısında güçlendirici nesne ışıklı dönen kırmızı kutu, diğer yarısında ışıklı dönen doldurulmuş tavuktu . .
Pedala ilk basış için gereken ortalama süre bu iki grup için aynıydı (Şekil 6.4, E). Her iki gruptaki kuşların yaklaşık %70'i , iki öğrenme deneyinden birinde aktif pedala art arda 10 basıştan 9'unu elde etti (Şekil 6.4, B). Gruplar, kritere ulaşma ortalama süresinde farklılık göstermedi ve her iki grupta da 9*

Resim: 6.4. Edimsel bir reaksiyonun geliştirilmesi. A. Pedala ilk basmanın ortalama gizli süreleri. B. Yüzde olarak ifade edilen, iki denemeden birinde öğrenme kriterine ulaşan kuş sayısı. Kuşlar yedi kuluçkadan elde edildi; her biri için öğrenme kriterine ulaşan civcivlerin yüzdesi hesaplandı. Bu yedi araçtan hesaplanan genel ortalamalar ve standart hatalar verilmiştir. B. İki öğrenme deneyinin her birinde kritere ulaşma süresi . İkinci deneydeki ortalama süre, hem sahte olarak çalıştırılan kontrol grubu hem de PMVH'si zarar görmüş tavuklar için birinciden önemli ölçüde daha kısaydı (P<0.005). (Johnson, Noth, 1986.)
ikinci deneyde kriterin elde edilmesi birinciden daha hızlıydı (Şekil 6.4, B). Ek olarak, her iki tavuk grubu da aktif pedalda harcanan süre açısından farklılık göstermedi. Böylece PMVH'si zarar görmüş civcivler, sahte kontrolden herhangi bir parametrede önemli ölçüde farklı olmadı ve her iki grup da yaklaşmaya çalıştıkları nesneyi görmek için pedala basmayı kolayca öğrendi.
Resim: 6.5. Edimsel yanıt geliştiren civcivler için tercih puanları. Takviye olarak kullanılan nesneler (dönen kırmızı kutu veya doldurulmuş tavuk) rayların uçlarına yerleştirildi (bkz. Şekil 3.2, B). Tercih göstergeleri formül kullanılarak hesaplandı
takviyelere yaklaşmaya
çalıştığı andaki platform hareket mesafesi)
Toplam platform hareket mesafesi
Sahte çalıştırılan kontrol civcivleri pekiştireç olarak gördükleri görsel nesneyi tercih ettiler ve ortalama tercih puanları %50'den önemli ölçüde fazlaydı. PMCH'si zarar görmüş kanatlılarda ortalama tercih indeksi, kontrol grubundan önemli ölçüde düşüktü (P<0.01) ve %50'den önemli ölçüde farklı değildi. (Johnson, Harp, 1986.)
platformda ve ardından teşvikler dahil. Civciv uyarıcıya yaklaşmaya çalıştığında çark döndü ve platform raylar boyunca hareket etti. Platformun 4 dakikada kat ettiği mesafeye bağlı olarak tercih indeksi hesaplandı (bkz. Şekil 6.5'in başlığı). Bu gösterge , testteki toplam aktivitenin yüzdesi olarak, tanıdık bir nesneye yönelik aktivitenin bir tahminini verdi . Sahte ameliyat edilen tavuklar , daha önce pekiştireç olarak kullanılmış bir nesneyi tercih ettiler; Hasarlı PMVG'ye sahip civcivler , yenisine kıyasla takviye nesnesi için daha belirgin bir tercih göstermeden şans düzeyinde hareket ettiler . Kontrol grubunun ortalama değerleri, deney grubununkinden önemli ölçüde yüksekti (Şekil 6.5).
Sahte grubun sonuçları , civcivlerin bir nesneyi görmek için pedala basmayı öğrenirken onun özelliklerini ezberledikleri gözlemini doğruladı (Bateson ve Reese, 1969). PMWG hasar gördüğünde, bu iki işlemin ayrışması gerçekleşir. Bu tür civcivlerde, nesne edimsel öğrenmede pekiştirici görevi görebilir, ancak tercih testinde tanıdık olarak tanınmaz . Bu rahatsızlığın doğasını anlamaya daha yakın olabilmek için , bir civcivin tanıdık bir nesneye seçici bir şekilde yaklaştığındaki davranışını analiz etmek ve ayrıca böyle bir durumda "tanıma" teriminin ne anlama gelebileceğini düşünmek gerekir .
Civciv, bir nesneye yaklaşmanın yanı sıra, zevkini ifade eden sesler çıkarabilir ve nesne ortadan kaybolduğunda alarm çığlıkları atabilir. Genellikle bu tür nesnelere baskı yapma literatüründe tanıdık denir. Aynı nedenle civcivin daha önce görmediği yeni bir nesneye yabancı denir. Baskının gücünü değerlendirmek için kullanılan test, tanıdık ve yeni bir nesnenin aynı anda maruz bırakılmasını içerir. Böyle bir tercih testi, bir tür tanıma testi olarak görülebilir (Sluckin, 1972). Hem insan (Baddeley, 1976) hem de hayvan (Gaffan, 1974, 1983) çalışmalarında, tanıdık ve tanıdık olmayan uyaranları ayırt etme yeteneğinin tanıma ve hafızaya bağlı olduğu düşünülmektedir. Bazı geçmiş deneyimlerin davranış üzerinde belirli bir etkisi olduğu için "hafızanın" bu sürece dahil olduğuna inanılmaktadır; "tanıma"dan söz edilir çünkü nesneler arasındaki ayrım, onlara aşinalık derecesine göre gerçekleşir.
bir tercih testinde, daha önce etki etmiş bir nesneye seçici olarak yaklaşamıyorsa, tanıma bozukluğu olduğunu varsayabilir miyiz ? Belki de öyle, ama hiç de gerekli değil. Bir nesneye seçici olarak yaklaşamama çeşitli şekillerde açıklanabilir ve ancak bunun için güvenilir gerekçeler varsa bir hafıza bozukluğundan söz edilebilir. Deneysel veriler, PMCH'si zarar görmüş civcivlerdeki davranış kusurlarının motor veya duyusal işlevlerdeki, uyanıklık düzeyi veya motivasyondaki ve diğer birçok faktördeki değişikliklerle açıklanmasının gerçeğe uymadığını göstermektedir (Bölüm 5.5, 7.4 ve 8.1). Bu, tavuk doğru pedala atladığında güçlendirici bir görsel uyaranın aydınlatıldığı ve döndürüldüğü yukarıda açıklanan deneylerle de doğrulanır . Takviye nesnesinin bu "aktivasyonu" ile kontrol tavukları ona yaklaşmak için pedaldan ayrıldı. PMVP'si zarar görmüş civcivler, istedikleri pedala kontroller kadar hızlı basmayı öğrenmekle kalmadılar, aynı zamanda pedaldan hızla çıktılar.Tercih testinde, kontrol
grubundan daha az aktif olmadılar, ancak bunun aksine, pedallarını yönlendirmediler. yeni bir nesne yerine pekiştirici bir nesneye doğru hareketler .
PMCH'si zarar görmüş civcivlerin daha önce gördükleri ve eğitimli kontrol civcivlerinin seçici olarak yaklaştığı bir nesneye seçici olarak yaklaşamamaları, PMCH'nin tanıma yeteneği ile doğrudan ilişkili olduğunun ön kanıtı olarak kabul edilebilir . Bu görüşü destekleyen ek kanıtlar sonraki bölümlerde verilmektedir. PMVD hasarı, damgalama sırasında belleğin ayrışmasına ve ilişkisel öğrenme için gereken belleğe yol açar.
Tanıma ve İlişkisel Öğrenme yavaş yavaş kayboldu, ancak farklı bir patolojinin belirtileri ortaya çıktı: X. M. , operasyon anından itibaren sürekli olarak devam eden şiddetli amnezi geliştirdi. Milner (1966), durumunun klinik tablosunu tanımladı ve annesinin sözlerinden anamnez verileri sağladı. X. M. çözümünde en ufak bir gelişme göstermeden aynı bilmeceyi her gün bir araya getirebiliyordu; içeriğine aşina olduğunu belli etmeden aynı dergiyi defalarca okuyabiliyordu. Benzer şekilde, ameliyattan sonra tanıştığı insanları, hatta yıllarca evini düzenli olarak ziyaret eden komşularını bile unutmuştu (Milner, 1966). Tanımadaki ciddi bozukluklara rağmen , "ayna çizme" gibi yeni bir motor beceri öğrenebildi. Ancak, bu görevi her tamamladığında, 5^?'ye sahip olduğunu inkar etti. al ve °; öğrenmeye bir "tanıdıklık" duygusu eşlik etmiyordu (Milner, 1962). Ona görsel ayrımcılık öğretilebilirdi . Kendisine sunulan görevlerden biri özellikle ilgi çekicidir, çünkü PMVP'si zarar görmüş tavuklar tarafından gerçekleştirilebilen görsel farklılaştırma görevleriyle bazı ortak özelliklere sahiptir (Bölüm 6.1). X. M.'den aynı anda sunulan bir daire ile elipsleri ayırt etmesi istendi (Sidman ve diğerleri, 1968). Figürler önündeki küçük panellere yansıtıldı . Üzerinde daire olan bir panele basarsa bir kuruş alırdı. Bu görevle başarıyla başa çıktı.
Orta-zamansal duygulanımlarda ortaya çıkana, semptomatolojide aynı olmasa da biraz benzeyen başka bir amnezi biçimi, bazen kronik alkoliklerde bulunur; bu türden bazı amnestik semptomlar, ilaç uygulamasıyla tersine çevrilebilir şekilde tetiklenebilir (Brown ve diğerleri, 1982, 1983). Kronik alkolizmle ilişkili sendrom ilk kez geçen yüzyılın sonunda Korsakov tarafından tanımlandı (bkz. Zangwill, 1983). Korsakoff sendromlu hastalarda beyin hasarı yaygın olabilir, ancak hipotalamustaki meme gövdelerindeki ve talamusun bazı bölümlerindeki patolojik değişiklikler değişmez bir özelliktir. Bu nedenle sendrom bazen diensefalik amnezi olarak adlandırılır (bkz. s. 283-284 ve bölüm 12.1.1). Korsakoff sendromunda , özellikle hatırlanacak nesne bağlam dışında sunulursa tanıma bozulur (Huppert ve Piercy , 1976). Ancak bu hastalarda dernekler oluşabilir . Bu ilk olarak Claparede tarafından tanımlanmıştır (Claparede, 1911, alıntılanan MacCurdy, 1928). Parmaklarının arasına iğne saklayarak Korsakov sendromlu bir hastanın eline batırdı. Bunu hatırlamadığını iddia etti, ancak Claparède eline tekrar yaklaştığında elini geri çekti. Görünüşe göre hasta, elinin hareketini bir iğne batmasıyla ilişkilendirdi, yani bir tür duyu-motor bağlantısı vardı; ancak neden elini geri çektiğini açıklayamadı.
hafıza bozukluğu olan iki hastada güvenilir kararlı koşullu tepkiler geliştirmeyi başardılar . Karmaşık bir işitsel ve görsel sinyal , koşullu bir uyaran görevi gördü. Ses kaynağı ve görsel uyaran - küçük bir sinyal ışığı - hastanın önünde, sol gözünün hemen altında bulunuyordu. Koşulsuz uyaran, kısa bir hava salvosuydu. Sağ göze yönelikti ve her seferinde yanıp sönen bir tepki ortaya çıkardı. Aparat çok dikkat çekici ve alışılmadık bir görünüme sahipti ve çalışma sırasında ve dinlenme molalarında görünür durumda kaldı. Bir hava salvosu ile birkaç ışık ve ses uyaran kombinasyonundan sonra, koşullu uyaran, pekiştirme iptal edildiğinde göz kırpmaya neden oldu. 10 dakika ila 24 saat arasındaki aralıklarla koşullu refleks yanıp sönmenin korunduğu gözlemlendi.Ancak denekler, eğitim prosedürünün koşullarına aşina görünmüyordu ve şartlandırılmış olarak kullanılan ışık ve zilin yanı sıra hava beslemesi için el aletini tanımıyorlardı. uyarıcı Bu sonuçlar çoğu amnezik hastanınkine çok yakındır. Nesnelerin veya olayların tanınmasını gerektiren görevlerde zayıf performans gösterirler (Gaffan, 1972; Warrington, 1974), görsel ayrım veya motor becerileri oldukça iyi öğrenme yeteneğine sahip olmalarına rağmen ( Talland, 1965; Sidman ve diğerleri, 1968; Gaffan, 1972). ; Savaş zil sesi, Weiskrantz, 1979). Tüm bu hastaların aynı klinik hafıza kaybı semptomları ve aynı beyin patolojisi ile karakterize olduğunu varsaymak yanlış olur (yukarıya bakın). Bununla birlikte, ortak semptomları belirlemeye çalışmak ve bu hastalarda benzer beyin hasarı olup olmadığını öğrenmek mantıksız değildir .
Hasta X. M.'de amnezinin bazı özellikleri tanımlandığında ve Korsakoff sendromunun ilk tanımlarına olan ilgi yeniden arttığında (Zangwill, 1966), araştırmacılar bu amnezilerin hayvan modellerini yaratmaya başladılar. Ancak, aramanın konusunun ne olduğunu açıkça anlamak çok önemlidir; Amnezik bir hastada bir uyarana aşina olduğunun sözlü olarak inkar edilmesi, hiçbir şekilde hayvanlarda amnezi üzerine çalışan bir deneycinin bekleyebileceği bir tepki değildir. Birkaç istisna dışında (Harlow ve Zimmerman, 1959), tanıma testi damgalama araştırması ve klinik nöropsikoloji dışında kullanılmamıştır . 1974'te Gaffan ( 1974) makaklarla deneyler için böyle bir test geliştirdiğinde ileriye doğru önemli bir adım atıldı. Test çok basitti. Maymuna yemek için üç delikli bir tabak gösterildi . "Basit" testte bir nesne sunuldu. Bir parça yiyecek içeren merkezi girintiyi kapladı. Maymun nesneyi hareket ettirip yemi çıkardıktan sonra maymun ile çanak arasına bir perde indirildi. Maymunun nesneye verdiği tepkiden 10 saniye sonra gerçekleştirilen karşılaştırma testinde tekrar sunuldu. Bu sefer, yem ortadakinin solunda veya sağında olacak şekilde girintinin üzerindeydi. Karşıdaki çöküntü, maymunun daha önce görmediği başka bir nesneyle örtülmüştü. Yeni nesne tarafından kapsanan bu çöküntü yiyecek içermiyordu. Hayvanın nesnelerden birini seçmesi ve daha önce gördüğü nesneyi hareket ettirerek yemi çıkarması sağlandı . Bu "örnek karşılaştırması" bir Tanıma testidir. Yeterli sayıda nesne varsa, bu tür örnekler birçok kez tekrarlanabilir. Görev, örneğin tanıdık bir nesneyle birlikte sunulan yeni nesnelerin sayısını artırarak daha da zorlaştırılabilir.
Bu ve diğer tanıma testleri (Mishkin ve Delacour, 1975), beyin hasarının hafıza üzerindeki etkisini incelemek için kullanılmıştır. Diensefalonun yapıları (Aggleton ve Mishkin, 1983a, b) veya temporal lobun medial kısmı (Zoia-Morgan ve Squire, 1984; Mahut ve Moss, 1985) hasar görmüştür. Yukarıda belirtildiği gibi, insan beyninin bu kısımlarına verilen hasar genellikle amneziye yol açar. Bu tür yaralanmalara sahip maymunlarda, nesne tanıma görevlerinin performansı bozulmuştur, ancak görsel farklılaşmanın geliştirilmesini gerektiren görevleri yerine getirme yeteneği çok daha büyük ölçüde korunmuştur. Bu nedenle, insanlar ve maymunlardaki spesifik beyin hasarının yanı sıra tavuklardaki PMVG hasarı benzer sonuçlara sahiptir : Tanıma için gerekli hafıza ile çağrışımsal öğrenme için gerekli hafıza arasında bir ayrışma vardır. Bu konular aşağıda bir kez daha tartışılacaktır ve PMGM'nin bellekteki rolü tartışılırken de dikkate alınacaktır (Bölüm 11.3).
PMVG'ye damgalamanın belirli bir bağımlılığı var mı?
Sec'de açıklanan deneylerin verileri. 6.1 ve 6.2, PMVP'nin işlevinin damgalamaya özgü olduğu sonucuna varmamızı sağlar . Böyle bir varsayımın doğruluğunu değerlendirmek için bir dizi faktörü hesaba katmak gerekir.
Etkili damgalama uyarıcısı olarak kullanılabilecek nesneler, damızlık kuşların civcivlerini çeşitli şekillerde etkiler. Bir sonraki tepkiyi ortaya çıkarmanın yanı sıra , bu tür uyaranlar bir güçlendirici görevi görebilir ve civcivlerin nesneyi daha fazla tanımasına olanak tanıyan sinir sisteminde değişikliklere neden olabilir. Böyle bir nesnenin aşağıdakilerin tepkisi üzerindeki etkisi doğrudan gözlemlenebilir. Bir pekiştirici olarak damgalayıcı bir uyaranın etkisi, deneysel verilerden tahmin edilebilir : pekiştiriciler karşılık gelen tepkileri güçlendirir ve bir damgalayıcı uyaran bu etkiyi ürettiği için (Bölüm 6.2), bu bir pekiştiricidir. Baskı yapan bir nesnenin sinir sisteminde bir tür "iz" bırakma yeteneği, kuşun önceden etkilenen nesneyi daha önce gözlemlenmemiş başka bir görsel nesneye tercih etmeye başlaması gerçeğinden kaynaklanır. Belirli bir nesneye damgalamanın başka bir sonucu, diğer görünür nesnelerden aktif olarak kaçınmaktır. Bu sonuç, elbette, doğrudan kuşların davranışlarını gözlemleyerek görülebilir.
Böylece, görsel deneyime sahip olmayan damızlık kuşların civcivlerine görsel nesnelerin maruz kalmasının etkilerini karakterize etmek için tamamen farklı yollar kullanılır. Bu, nöral mekanizmaların daha fazla analizi için iyidir, çünkü mantıksal olarak birkaç farklı sinir sürecinin katılımını hayal etmek mümkündür. Dolayısıyla, damgalayıcı uyaran: 1) bir yaklaşma tepkisine neden olur; 2) takviye görevi görür ; 3) sinir sistemlerinde bir tahriş izi oluşumuna yol açan süreçlere neden olur; 4) diğer göze çarpan nesnelerden kaçınmaya yol açabilir .
Görsel deneyimi olmayan kuşlarda bir yaklaşma tepkisini uyandırarak , baskı yapan uyaran Pavlov'a göre koşulsuz bir uyaran, yani özel bir ayrıntıya girmeden bir yanıt veya bir dizi tepki ortaya çıkaran bir uyaran gibi davranır (Mackintosh, 1974). Baskı yapan nesneler edimsel tepkileri pekiştirirken, pekiştirici görevi görürler. Baskı yapan nesnelerin pekiştirici özellikleri, civcivin bu nesnelere verdiği tepkileri etkiler mi ? Hoffman ve diğerlerinin ( 1972) deneyleri durumun böyle olduğunu göstermektedir. Bu araştırmacılar, ördek yavrularının kendilerine hareketli bir uyaran sunulur sunulmaz alarm vermeyi bıraktıklarını gösterdi . Uyaran hareketsiz ise, alarm çağrıları devam eder. Hoffman ve arkadaşları ( 1972) deneyi , durağan bir uyaranın sunumu ile hareket halindeki sunumunun birbirini izleyeceği şekilde kurdu. Bu koşullar altında, durağan uyaran aynı zamanda kaygı ile ilişkili seslendirmeyi bastırma yeteneği de kazanmıştır . Bu sonuç, hareketli bir uyaranın pekiştirici özelliklerinin, alarm çığlıklarının bastırılmasına koşulsuz olarak refleks olarak bir tepki ortaya çıkararak, uyaranın diğer özelliklerine aktarıldığını gösterir. Hoffman ve Ratner (1973), damgalayıcı bir uyaranın pekiştirici yönlerinin, o uyaranın diğer özelliklerini hatırlamayı kolaylaştırdığı genel fikrini geliştirdiler.
Örnek olarak basit bir model kullanarak damgalamada yer alan bazı nöral süreçleri ve bunların olası etkileşimlerini açıklamak yararlıdır. Böyle bir modelin şeması, Şek. 6.6A , Civciv, etkili bir yapay baskı nesnesi olan uyaran I ile sunulur . Tüm görsel uyaranlar bu davranışı ortaya çıkarmaz; örneğin, darı büyüklüğündeki uyaranlar bir gagalama tepkisine neden olur. Buna dayanarak, duyusal girdi ile yaklaşma tepkisini düzenleyen motor sistem arasında bir tür filtrenin varlığı hakkında bir varsayımda bulunulabilir. Böyle bir filtre, Şekil l'de F harfi ile gösterilir. 6.6, L, ancak basit olması için diyagramların geri kalanından çıkarılmıştır. Modele göre, "I" uyarısı birkaç nöral yolu aktive eder. Bu yollardan birindeki uyarılmış aktivite, yaklaşımı düzenleyen nöronları (Şekil 6.6, Ek) zayıf bir şekilde uyarır - koşulsuz bir refleks reaksiyonu. Bu yaklaşım belirli bir konuya özgü değildir.
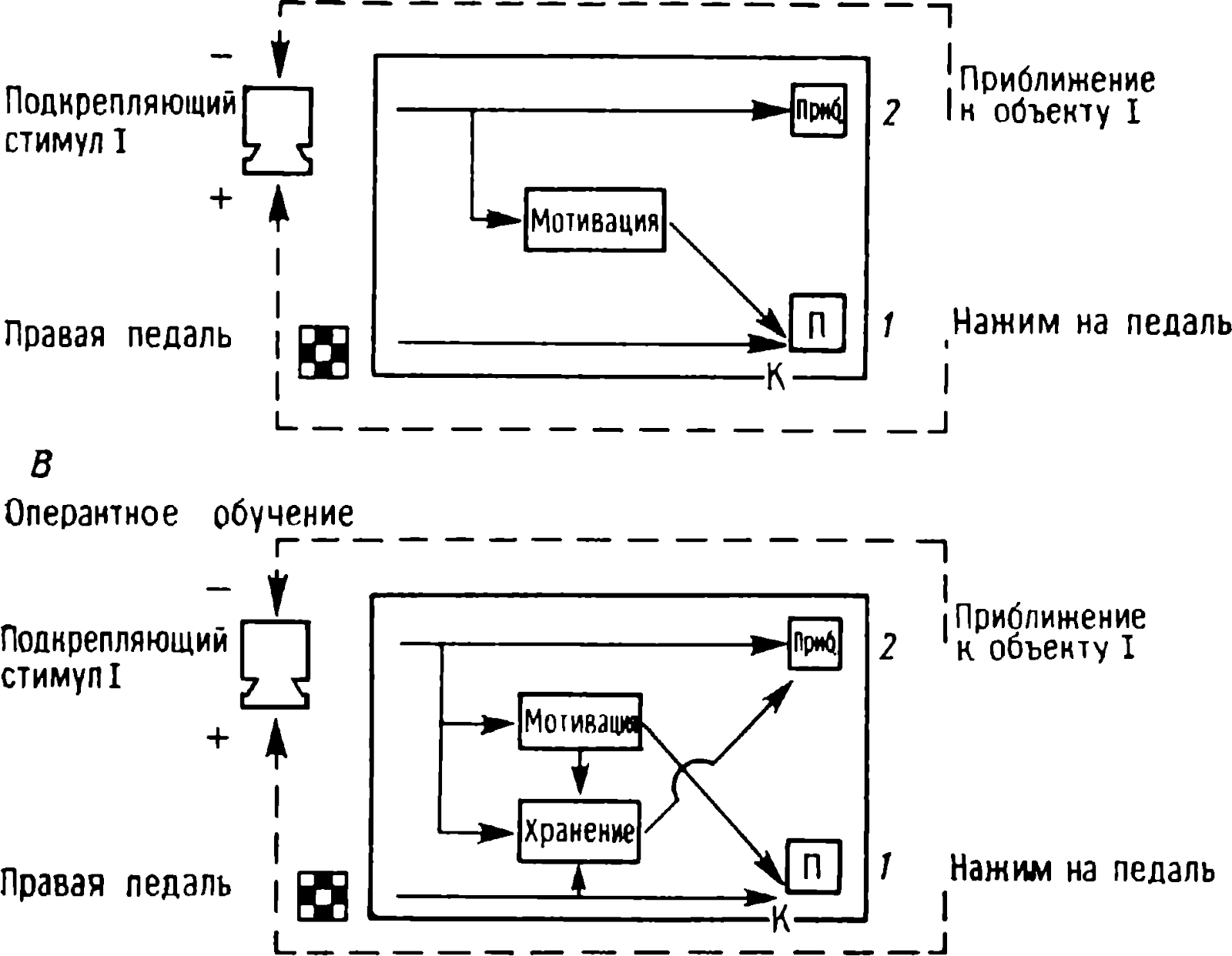
Resim: 6.6. Eğitim sırasında meydana gelen bazı süreçlerin şeması. Büyük dikdörtgen, civcivin merkezi sinir sistemini temsil eder. İçerideki dikdörtgenler , belirli işlevleri olan sinir yapılarını gösterir . Büyük kutudaki oklar , civciv beyninde meydana gelen ve muhtemelen multisinaptik yolların aracılık ettiği etkileşimleri gösterir ; dışındaki oklar , civcivin davranışının sonuçlarını gösterir. İki kırmızı ışıklı yüzeye sahip dönen bir kutu olan Uyaran I, etkili bir yapay damgalama nesnesidir. A. Bir damgalama durumunda Uyaran I'e maruz kalma sırasında meydana gelebilecek olaylar . F, yalnızca bu diyagramda gösterilen ve tüm görsel uyaranların bir yaklaşma tepkisi uyandırmadığını vurgulamak için tanıtılan bir filtredir. yaklaşık yaklaşımı kontrol eden sinir sistemi anlamına gelir. B. Civciv edimsel öğrenme aparatındadır ( Şekil 6.3). İçinde hareket ederken sağda duruyor, cihazın zemininde aktif pedal. Bu pedal , resimde gösterildiği gibi kareli bir desenle kaplanmıştır. R, sağ pedala yaklaşma ve üzerine atlama tepkisini düzenleyen bir nöron sistemidir . Bunun sonucu, uyaran I'in dahil edilmesidir (+). Göründüğünde, civciv ona yaklaşır (2) ve pedaldan ayrılır. Sonuç olarak, uyaran I kapatılır (-). A ve B'de gösterilen süreçler edimsel öğrenmede eşzamanlı olarak gerçekleşir ve Diyagram C'de birlikte gösterilir. Daha ayrıntılı açıklamalar için metne bakın;
nesne. Baskı yapan uyaran, pekiştirici ile aynı şekilde hareket eder ve model, bu eylemin motivasyon sistemi tarafından aracılık edildiğini varsayar (ayrıca bkz. Bölüm 9). Uyaran I tarafından uyarılan nöral yollardan biri, hafıza izlerinin depolanmasına yol açar. Maruz kalma süresi boyunca , uyaranın nöral temsili yavaş yavaş bu depolamada oluşur. Bu temsilin oluşması için gerekli koşul , motivasyon sisteminden bir girdinin varlığıdır. İz oluştukça iz deposundan gelen çıkış sinyali artar ve bunun sonucunda civcivin I uyaranına yaklaşması artar. Civcivlerin yeni nesnelerden kaçınmaya başladığı süreçler modelde gösterilmemiştir. Kaçınma davranışı, anne ikamesi olan belirli bir nesneye bağlanmanın bir sonucudur: civciv bu nesneye daha aşina hale geldikçe, ondan keskin bir şekilde farklı olan diğer nesnelerden kaçınmaya başlar. Uyaran I'in nöral modelinin oluşumunda , tüm I olmayan nesnelerden kaçınıldığı görülmektedir (bkz. Şekil 7.10 ve 7.11). Kaçınma davranışının gelişimi, karşılaştırma sağlayan bir sinir sistemine dayanabilir: saklanan bilgi ile sinir sisteminde yeni bir nesne tarafından uyarılan aktivite arasındaki güçlü uyumsuzluk, kaçınma sisteminin aktivasyonuna yol açar. Bununla birlikte, böyle bir karşılaştırma sistemi söz konusu olsa bile, bu konu birkaç yıldır analiz konusu olmasına rağmen, etki mekanizması hakkında çok az şey bilinmektedir (Horn, 1952, 1967; Sokolov, 1960; Vinogradova, 1970; Palm, 1982; bkz. Şekil 1.12).
Şek. 6.6 , Johnson ve Horne'un (Bölüm 6.2) deneylerinde geliştirilen edimsel reaksiyonun performansına dahil olan sinir süreçlerini basitleştirilmiş bir biçimde ele almak için de kullanılabilir . Eğitim aparatının zeminindeki sağ pedal, onu sol pedaldan ayıran parlak kareli bir desenle kaplanmıştır (Şekil 6.3). Civciv odanın etrafında az çok rastgele hareket ettiğini varsayalım. Şans eseri, örneğin kameranın sağ tarafındaki "aktif" pedala basıyor. Duyusal yollarda, sağ "damalı" pedalın etkisi altında aktivite ortaya çıkacaktır . Bu aktivite, tavuğun hareketini kontrol eden, sağ pedala giden ve üzerinde duran nöron sisteminin uyarılmasıyla çakışır (Şekil 6.6, B, kare P). Civciv pedal üzerinde durduğunda, uyaran I açılır.Bu uyaranın uyandırdığı motivasyonun etkisi, duyusal yollardaki ve nöronal sistem P'deki (Hebb, 1949 ) ortak aktivite sırasında K temasındaki (Şekil 6.6b ) iletimi kolaylaştırabilir. ). Uyaran I açıldığında, bir yaklaşma tepkisi ortaya çıkarır. Bu durumda civciv pedalı bırakır ve uyaran I kapatılır. Lenok civciv odanın zemininde yürümeye başlar. Sol pedala basarsa , uyaran I açılmaz. Sağ pedala basarsa, uyaran I'in güçlendirici etkisi yine K temasında rahatlamaya yol açar. İşlemin tekrarlanmasıyla bu bağlantı kademeli olarak güçlenir ve sağ pedaldaki kontrol deseni sağ pedala basma sisteminin kontrolünü artırır. Doğru pedala basma becerisi oluşur. Bu alışkanlığın nöral mekanizması, K temasında iletimi kolaylaştırdığı varsayılan mekanizmadır (Şekil 6.6, 5, C) .Bu mekanizmanın, bir alışkanlığın oluşumunun altında yatan pek çok mekanizmadan yalnızca biri olduğunu vurgulamak önemlidir . İlişkisel öğrenmede "hafıza" periferiktir, iletimin kolaylaştırıldığı temasta oluşur (K). Modelde, bu bellek, Şekil 2'de gösterilen merkezi izleme havuzunda özel olarak temsil edilmemiştir. 6.6.
Uyaran I'in aktivasyon döneminde, uyandırdığı sinyaller iz depolama ve motivasyon sistemlerine geçer, böylece hafızada bir sinyal izi oluşur (Şekil 6.6, K). Tüm edimsel öğrenme dönemi boyunca civciv beyninde meydana gelen süreçler ve etkileşimler şematik olarak Şekil 1'de özet halinde gösterilmiştir. 6.6B Civciv edimsel öğrenmeden sonra bir tercih testi yapıldığında, uyaran I'e yaklaşır ve onu tanır çünkü o uyaranın izi hafızadadır. Model, işlemsel öğrenmeden önce bellek deposunu silmenin ana sonuçlarını açıklar . Bu deneysel manipülasyonun bir sonucu olarak, Şekil 2'de gösterilen işlemler, 6.6, B, Şek. 6.6, B, burada bellek depolama gösterilmez. Bu koşullar altında, civciv edimsel bir tepkiyi öğrenme konusunda oldukça yeteneklidir, ancak pekiştiriciye aynı görsel çekiciliğe sahip diğer herhangi bir nesneden daha fazla yaklaşmayacaktır. Tabii ki, bu şema aşırı derecede basittir, çünkü sağlam bir civcivin edimsel öğrenme ilerledikçe uyaran I'e yaklaşma eğiliminin artması beklenmelidir. Sonuç olarak, civciv pedallara daha az zaman harcamak zorunda kalır ve sonuç daha az takviye olur. Bu nedenle, tavuğun bir şekilde çelişkili dürtülerini optimize etmesi gerekiyor.
Şekil 1'de gösterilen çeşitli işlemlerden hangisi. 6.6, eğitimden önce veya kısa bir süre sonra PMVP'de iki taraflı hasar olması durumunda ihlal edilebilir mi? Bu, motivasyon sistemini yok ederse, o zaman bellekte uyaran I'in izi oluşmayacak ve tavuk bunu tanımayacaktır (Bölüm 5.2 ve 5.3). Ayrıca bu uyarıcı pekiştireç olarak kullanılamaz (Şekil 6.6, B). Sonuç olarak, bu tür hasara sahip civcivler edimsel beceriyi öğrenemeyeceklerdir. Hâlâ öğrendikleri için, PMVP'deki hasarın motivasyon sistemini yakalamadığı sonucuna varabiliriz. PMCG'deki hasar, damgalayıcı bir uyaranın koşulsuz bir uyaran olarak bir yaklaşma tepkisi ortaya çıkarma yeteneğini etkiler mi? Açıkçası hayır, çünkü bu tür yaralanmalara sahip civcivler görsel nesnelere yaklaşıyor. Sonuç olarak, PMVP'nin kaldırılması, Şekil 1'de gösterilenin ihlali ile ilişkili değildir. 6.7A, uyaran I tarafından uyarılan sinyalleri doğrudan yaklaşma sistemine ileten bir yol.
Baskılama prosedüründen önce PMVG'nin iki taraflı olarak çıkarılmasıyla, yaklaşma tepkisi korunsa da öğrenme sürecinde kontrol tavuklarında olduğu gibi artmaz (Bölüm 5.1 ve Şekil 5.2). Bu sonuçlar, motivasyon sisteminin, yaklaşma sistemine giden yolda doğrudan davranışın kolaylaştırılmasını etkilemediğini düşünmemizi sağlar (Şekil 6.6, K). PMVP'si zarar görmüş tavuklar, ne damgalama durumunda ne de edimsel beceriyi öğrenirken kendilerine sunulan I uyaranını tanımazlar. Böyle bir etki ancak bellek izlerinin depolanmasını yok ederek elde edilebilir. Bu teşhir , görsel nesnenin bir yaklaşma tepkisi ortaya çıkarma veya bir güçlendirici olarak hareket etme yeteneğini bozmaz. Buradan, model tarafından gerçekleştirilen tüm işlemler arasında SMVG'nin büyük olasılıkla izleri saklayacağı sonucu çıkar. Bu departman ya kendi başına bir izleme havuzu işlevi görebilir ya da başka bir yerde depolamaya erişim sağlayabilir. Yukarıda, ikinci varsayımın deneysel verilerle çeliştiğini gösteren argümanlar sunduk (Bölüm 5.5).
Kossut ve Rose ( 1984) , tavuklarda çağrışımsal öğrenmenin , radyoaktif 2-deoksiglikozun ventral hiperstriatumun medial kısmına dahil edilmesini arttırdığını bulmuşlardır. Bu bölge, PMWG ile aynı görünmektedir. Kossuth ve Rose pasif kaçınma eğitimi kullandılar (Lee-Teng ve Sherman, 1966; Cherkin, 1969). Bu prosedürde, küçük bir boncuk (2-4 mm çapında), insanlar için çok hoş olmayan bir tada sahip bir madde olan metil antranilat içine daldırılır . Tavuklar böyle bir boncuğu gagaladıklarında, gözlerini kapasalar da, gagalarını açsalar da, başlarını sallasalar da çoğu iğrenme belirtileri gösteriyordu. Birkaç saat sonra aynı kuru boncuk teklif edildiyse , çoğu onu gagalamadı. Daha önce suya batırılmış bir boncuğu gagalayan kontrol civcivleri bundan kaçınmadı.
Metil antranilat batırılmış mürveri gagalayan civcivler için bu, tatsız bir deneyimin uyarısı haline geldi , yani civcivlerin boncuğu gagalaması ile bu eyleme eşlik eden tatsız deneyim arasında bir bağlantı oluşturduğu sonucu çıktı. Bu nedenle, pasif kaçınma ilişkisel öğrenmeyi ifade eder. Bu tür bir eğitim , ilk bakışta Bölüm 1'de açıklanan sonuçlarla çelişiyor gibi görünen PMWG'nin (Kossut ve Rose, 1984; Stewart ve diğerleri, 1984) katılımını gerektirir . 6.1 ve 6.2, PMVP'ye verilen hasarın ilişkisel öğrenme üzerindeki etkisinin olmaması hakkında. İlişkisel öğrenme, PMCH'nin yokluğunda devam ederse, o zaman, elbette, böyle bir öğrenmeden sonra beynin bu bölgesindeki hücre döngüsündeki değişiklik şaşırtıcıdır.
Bölümde açıklanan deneylerde. 6.1 ve 6.2'de, takviye pozitifti, ancak metil antranilat ile yapılan deneylerde negatifti . Civcivler, bu madde ile kaplanmış bir boncuğu gagalamanın bir sonucu olarak, eğitimden önce var olan reaksiyonu bastırdılar: parlak küçük nesneleri gagalama eğilimleri azaldı. Bu davranış, olanların özünü açıklayabilir. Gaffan ve diğerleri ( 1984) , forniks transeksiyonunun maymunlarda çağrışımsal öğrenme üzerindeki etkisini ayrıntılı olarak inceledi . Fornix, diensefalondaki diğer yapılara çıkıntı yapan hipokampal hücrelerden gelen aksonları içerir. Forniks hasarından sonra tanıma yeteneği bozulur ( Gaffan , 1974; Gaffan ve Weiskrantz, 1980). Bunun tersine, bazı şartlandırılmış reflekslerin performansı için gerekli olan hafıza korunur (Gaffan, 1974). Bu ifadeden, zararın çağrışımsal öğrenmenin bazı biçimlerini etkilemediği sonucu çıkar. Bundan tam olarak ne etkilenir ve ne korunur? Gaffan ve diğerleri ( 1984), çok sayıda deney serisinden elde edilen verilere dayanarak, forniksi kesmenin yiyecek aramak gibi “doğal” davranışsal eğilimleri içeren çağrışımsal görevlerin performansını bozmadığı sonucuna varmışlardır ; ancak, gelişimi hayvanın "doğal eğilimine" aykırıysa, reaksiyonun yürütülmesi bozulur (Douglas, 1967). Tavuklarda, PMVP'nin yokluğunda başarıyla çözülen her iki ilişkisel görev de "doğal" reaksiyonlara dayanıyordu. Güçlendirici olarak ısının kullanıldığı testte, ödül olarak bir miktar ılık hava verildi (Bölüm 6.1). Deneyin yapıldığı oda soğuktu ve eğitilen civcivler test sırasında ısınmaya çalıştı. Sec'de açıklanan deneyde. Şekil 6.2'de (Johnson ve Horn 1984, 1986), pekiştirici uyaran civcivlerin herhangi bir eğitim almadan yaklaştıkları bir nesnenin görüntüsüydü. Bu iki deneyimin aksine, pasif kaçınma, küçük, parlak nesneleri gagalamaya yönelik güçlü bir eğilimin bastırılmasını gerektirir; bu durumda, edinilen davranış civcivlerin gagalama eğilimleriyle tutarsızdı. Bu analiz, en azından , bir yandan primatlardaki hipokampus-fornix sisteminin ve diğer yandan tavuklardaki PMVG sisteminin tanımadan sorumlu olduğunu ve belirli ilişkisel öğrenme türlerinde yer aldığını gösteriyor. Aynı zamanda, deneysel veriler, bu sistemlerin tüm ilişkisel öğrenme biçimlerine katılma olasılığını dışlamaz.
Bir hayvan, bir sinyal ile bir pekiştireç arasında bir bağlantı geliştirdiğinde, iki şey olabilir. Bir becerinin kazanılması , uyaranın ateşlediği alıcılar ile tepkiyi kontrol eden nöronlar arasındaki bağlantıların güçlendirilmesinden biraz daha fazlasını gerektirir (ayrıca bkz. Bölüm 12.1.1). Dahası, normal koşullar altında hayvan , belirli bir tepkiyle ilişkilendirdiği uyaranı tanıyormuş gibi görünür, tıpkı normal bir kişinin aynı anda Claparède'in elinin yaklaşmasından kaçınıp Claparède'in iğneyi parmaklarının arasına saklayan adam olduğunu anlaması gibi. Claparède tarafından tanımlanan amnezik hastalarda , elin geri çekilmesi için gerekli olan hafıza , tanıma için gerekli olan hafızadan ayrılmıştır . Normal insanlarda, davranışın bu iki bileşeninin altında yatan bellek sistemlerinde büyük olasılıkla nöronal değişiklikler meydana gelir. Buradan hareketle çağrışımsal öğrenmede bu iki bellek sisteminin paralel çalıştığı söylenebilir. Bu tam olarak , örneğin (Şekil 6.6, B), pedalın damalı deseninin neden olduğu sinyaller izlerin merkezi deposuna kaydedilirse ve aynı zamanda afferentler arasındaki K kontağından geçerken yükselirse tam olarak ne olur? girişi ve “sağ pedal basma sistemi”. Bu ifade, Şekil 1'de gösterilmektedir. 6.7. Periferik bellek, sensorimotor yolunda yer alan K kontağında artan iletim ile temsil edilir . "Depolama" etiketli kutu , pedal üzerindeki dama tahtası deseninin özelliklerini temsil eden merkezi bellek sistemine* karşılık gelir . Böyle bir organizasyonla, duyu-motor yol kesildiğinde bozulacak olan alışkanlık ortadan kalkmadan izlerin merkezi deposu yok edilebilir . Bu varsayımı doğrulayan deneysel veriler var. İlk olarak Benowitz ( 1972) , PMSH'yi etkileyen lezyonların civcivlerde yukarıda açıklanan pasif kaçınma alışkanlığını bozmadığını bulmuştur; tavuklar yapmaya devam ediyor. Eğer bu doğruysa ve beynin bu bölgesindeki (Kossut ve Rose, 1984; Stewart ve diğerleri, 1984) biyokimyasal değişiklikler, pasif kaçınma öğrenmesi sırasında izlerin kaydedilmesine bu bölgenin katılımını yansıtıyorsa , o zaman bu kayıt gitmelidir. yürütme becerisini destekleyen izlerin oluşumuna veya pasif kaçınma tepkisinin sensorimotor bileşenine paralel olarak. İkincisi, beynin yanal bölgesindeki hasar, öğrenme izlerinin korunmasını büyük ölçüde bozar.
Yu-140
Operasyon eğitimi Tavuk
pasif kaçınma (Benowitz, 1972); bu hasar, baskı sırasında elde edilen tercihin korunmasını etkilemez (Bölüm 5.2).
Klasik koşullu reflekslerle ilgili bazı çalışmalarda elde edilen görünüşte paradoksal sonuçlar , iki paralel işletim belleği sisteminin varlığı varsayılarak da açıklanabilir. Bir tavşan veya kedi gözünün korneasına yöneltilen bir hava patlaması , üçüncü göz kapağının yanıp sönmesine ve kasılmasına neden olur. Bir hava patlamasını bir ses tonuyla yeterince birleştirirseniz, izole edilen ton bu tepkiyi uyandırmaya başlayacaktır. Böyle bir koşullu refleks , hipokampus da dahil olmak üzere, havlamayan kedi ve tavşanlarda geliştirilebilir (Thompson ve diğerleri, 1983). Bununla birlikte, öğrenme sırasında, hipokampal nöronların aktivitesi oldukça spesifik bir şekilde değişir (Berger ve Thompson, 1978). Bu verilerin birleştirilmesi, hipokampüsün öğrenme sürecine dahil olduğunu, ancak şartlandırılmış refleksin sensorimotor bileşeninin oluşumu için gerekli olmadığını göstermektedir. Yukarıdaki değerlendirmelerde gösterildiği gibi, bu bileşen, tonla etkinleştirilen reseptörleri ve yanıp sönen refleksi kontrol eden nöronları bağlayan temasların güçlendirilmesinden oluşabilir. Ton özellikleri hakkındaki bilgiler, hipokampusu da içeren merkezi hafıza sisteminde (Şekil 6.7) de saklanabilir.
Kuş beynine geri dönersek, PMVP'nin her zaman tanımaya dahil olması durumunda, hafızanın damgalama veya diğer öğrenme biçimleri sırasında oluşmasından bağımsız olarak, bunun ona bazı genel işlevler atfetmemize izin vereceği söylenebilir. Bunu kanıtladıktan sonra, PMSH sisteminin bellek işlemleri için genel bir öneme sahip olduğu düşünülebilir. Kaydedilen bilgiler sadece belirli nesnelerin izlerinden oluşmamalı, aynı zamanda bu izlerin hayvanın bu nesnelere verdiği tepkilerin sonuçlarına ilişkin (örneğin küçük bir boncuğun iğrenme yaratıp yaratmadığı gibi) belirli kurallardan oluşmalıdır. Tadı hoş olmayan bir boncuğu gagalayan bir kuş, çok farklı koşullar altında karşılaşabileceği benzer boncukları artık gagalamayabilir (Dickinson, 1980, özellikle bölüm 3). Bu varsayımlar daha ayrıntılı olarak ele alınacaktır (Bölüm 7.3, 11.3 ve 12.1.1). Bununla birlikte, PMCG'nin ilişkisel öğrenmedeki rolünü anlamak için birçok ek veriye ihtiyaç olduğu açıktır. Aynı zamanda, halihazırda mevcut verilere dayanarak, PMVP'nin sadece baskıda değil, aynı zamanda baskıda da yer aldığı varsayılabilir. diğer öğrenme süreçleri.
Özet
Görsel nesneler: 1) eğitimsiz civcivlerde bir yaklaşma tepkisini uyandıran koşulsuz uyaranlar olarak; 2) takviye olarak; 3) nesnelerin nöral modellerinin oluşumuna yol açan süreçlerin tetikleyicisi olarak (tanıma hafızası). Bölüm , PMVP'deki iki taraflı hasarın fonksiyon 1 ve 2'yi bozmadığını gösteren veriler sunar . Bu hasarlar, nesnenin (3) tanınmasına izin vermez, ancak bazı ilişkisel öğrenme türlerini etkilemez. Tanıma ve diğer ilişkisel öğrenme biçimlerinde benzer bellek ayrışmasını gösteren insanlardan ve diğer primatlardan elde edilen verilerin bir incelemesi verilmiştir .
EDEBİYAT
Aggleton J. R., Mishkin M. Maymunlarda medial talamik lezyonları takiben görsel tanıma bozukluğu, Neuropsychologia, 1983a, 21, 189-197 .
Aggleton JP, Mishkin M. Maymunlarda sınırlı mediat talamik lezyonları takiben hafıza bozuklukları, Exp. Brain Res., 1983b, 52, 199-209.
Bateson PPG, Reese EP Baskı durumunda göze çarpan uyaranların pekiştirici özellikleri, Anim. Behav., 1969, 17, 692-699.
Baddeley AD Hafıza psikolojisi. Harper ve Row, New York, 1976. oenowitz L. Ön beyin ablasyonlarının civcivlerde kaçınma öğrenmesi üzerindeki etkisi. Fizyol. Behav., 1972, 9, 601-608.
Berger TW, Thompson RF Tavşan niktitatif membran tepkisinin klasik koşullanması sırasında limbik sistemdeki nöronal plastisite. I. Hipokampus. beyin. Res., 1978, 145, 323-346.
orierley J, B. Amnezik Durumların Nöropatolojisi. İçinde: Amnesia (2. baskı) (editörler / W.M. Whittey ve O.L. Zangwill), 1977, s. 199-243, Butterworth. Londra.
on*
Brown J., Lewis V., Brown M., Horn G., Bowes J. B. İki ilacın (diazepam ve lorazepam) neden olduğu geçici amneziler ile organik kaynaklı amnezi arasında bir karşılaştırma. Nöropsikoloji, 1982, 20, 55-70.
Campbell BA, Pickleman LR . Güçlendirici bir uyaran olarak baskı yapan nesne, J. comp. fizik. Psychol., 1961, 54, 592-596.
Cherkin A. Bellek konsolidasyonunun kinetiği ve amnezik tedavi parametrelerinin rolü, Proc. nat. Acad. bilim ABD, 1969, 63, 1094-1101.
Claparede E. Recognition ve moiite. Archs Psychol., Cenevre, 1911, 11, 79-90.
Dickinson A. Çağdaş hayvan öğrenme teorisi, 1980, Cambridge University Press, Cambridge.
Douglas RT Hipokampus ve davranış, Psychol. Bull., 1967, 67, 416-442.
Gaffan D. Forniks lezyonları olan sıçanlarda tanıma hafızası kaybı. Nöropsikoloji, 1972, 10, 327-341.
Gaffan D. Forniksin kesilmesinden sonra maymunların hafızasında tanıma bozukluğu ve bozulmamış çağrışım, J. comp. fizik. Psychol., 1974, 86, 1100-1109.
Gaffan D. Hayvan amnezisi: bazı kopukluk sendromları? İçinde: Hipokampusun Nörobiyolojisi (ed. W. Seifert), 1983, s. 513-528, Academic Press, New York.
Gaffan D., Weiskrantz L. Maymunlar tarafından rasgele yemlenmiş numunelerle gecikmeli eşleşmemede yenilik etkileri ve lezyon etkileri, Brain Res., 1980, 196, 373-386.
Gaffan D., Saunders RC, Gaffan EA, Harrison S., Shields C., Owen MJ Maymunlarda çağrışımsal hafıza üzerine forniks transeksiyonunun etkileri: öğrenilmiş eylemde hipokampusun rolü, Quart. J. exp. Psychol., 1984, 36, 173-221.
Harlow HF, Zimmerman RR Bebek maymunda duygusal tepkiler, Science, 1959, 130, 421-432.
Hebb DO Davranış organizasyonu. John Wiley, New York, 1949.
Hoffman HS, Ratner AM Sabit bir baskılayıcı uyaran olan Psychon tarafından davranış kontrolünün pekiştirilmiş bir modeli. Rev., 1973, 80, 527-544.
Hoffman HS, Schiff D., Adams J., Searle JL Seçici pekiştirme yoluyla gelişmiş sıkıntı seslendirmesi, Science, 1966, 151, 352-354.
Hoffman HS, Eiserer LA, Singer D. Durağan bir damgalayıcı uyaran, Psychon ile davranış kontrolünün kazanılması. Sci., 1972, 26, 146-148.
Horn G. Düşüncenin nörolojik temeli, Deniz Kızı, 1952, 18, 17-25.
Horn G. Nöronal alışma mekanizmaları, Nature, Londra., 1967, 215, 707-711.
Boynuz G. Beyinde bilgi depolama. Evcil civcivde damgalama çalışması. İçinde: Omurgalı nöroetolojisindeki gelişmeler (eds. J.-P. Ewert, RR Capranica ve DJ Ingle), 1983, s. 511-541, Plenum Press, New York.
Huppert FA, Piercy M. Amnezik hastalarda tanıma belleği: zamansal bağlamın etkisi ve malzemenin aşinalığı, Cortex, 1976, 12, 3-20.
Johnson MH, Horn G. Beyin lezyonlarının damgalama ve ilişkisel öğrenme görevi üzerindeki farklı etkileri, Neurosci. Letonya İlave, 1984, 18, 131.
Johnson MH, Horn G. Civciv ön beyninin kısıtlı bir lezyonu ile tanıma belleği ve çağrışımsal öğrenmenin ayrışması, Neuropsychologia, 1986, 24, 329-340.
Kossut M., Rose SPR Pasif kaçınma eğitimi sırasında civciv beyin yapılarına Diferansiyel 2-deoksiglikoz alımı. Nörobilim, 1984, 12, 971-977.
Lee-Teng E., Sherman SM Civcivlerde tek deneme öğreniminin hafıza pekiştirmesi. Proc. nat. Acad. bilim ABD, 1966, 56, 926-931.
McCabe BJ, Cipolla-Heto J., Horn G „ Bateson PPG Damgalamadan sonra civcivin hiperstriatum ventralinde yer alan bilateral lezyonların amnezik etkileri, Exp. Brain Res., 1982, 48, 13-21.
MacCurdy JT Psikoloji ve fizyolojide ortak ilkeler. Cambridge
University Press, Londra, 1928.
Vlackintosh NJ Hayvan öğreniminin psikolojisi. Academic Press, Londra, 1974.
Mahut H., Moss M. Belleğin pekiştirilmesi - hipokampus yeniden ziyaret edildi. İçinde: Hafızanın nöropsikolojisi (editörler N. Butters ve L. Squire), 1985, s. 297-315, Guilford Press, New York.
Mair WGP, Warrington EK, Weiskrantz L. Korsakoff'un psikozunda hafıza bozukluğu. İki olgunun nöropatolojik ve nöropsikolojik incelemesi, Brain, 1979, 102, 749-783.
Milner B. İki taraflı hipokamp lezyonlarına eşlik eden hafıza sorunları. İçinde: Physiologie de 1'hippocampe, Colloques Internationaux, 1962, no. 107, s. 257-272, Paris: CNRS.
Temporal loblarda ameliyatı takiben Milner B. Amnezi. İçinde: Amnezi. (der. CWM Whitty ve OL Zangwill), 1966, s. 109-133, Butterworth, Londra.
Mishkin M., Delacour J. Maymundaki kısa süreli görsel hafızanın analizi, J. exp. Psikol.: Animasyon. davranış. Süreç, 1975, 1, 326-334.
Palm G. Sinir meclisleri. Springer, W. Berlin, 1982.
Peterson N. Damgalanmış bir uyaranın sunumuyla davranışın kontrolü, Science, 1960, 132, 1395-1396.
Scoville WB, Milner B. Bilateral hipokampal lezyonlardan sonra son hafıza kaybı, J. Neurol. beyin cerrahisi. Psikiyat., 1957, 20, 11-21.
Sidman M., Stoddard LT, Mohr JP Bilateral hipokampal lezyonları olan bir hastada anlık belleğin bazı ek nicel gözlemleri, Neuropsychology, 1968, 6, 245-254.
Sluckin W. Damgalama ve erken öğrenme. Methuen, 1972, Londra.
Sokolov TR Nöronal modeller ve yönlendirme refleksi. İçinde: Merkezi sinir sistemi ve davranış (ed. M. A. Brazier), 1960, s. 187-276. Josiah Macey Jr. Vakıf, New York.
Stewart MG, Rose SPR, King TS, Gabbot PLA, Bourne R. Pasif kaçınma eğitiminin ardından civciv medial hyperstriatum ventrale'deki sinapsların hemisferik asimetrisi: stereolojik bir araştırma, Devi. Brain Res., 1984, 12, 261-269.
Talland GA Düzensiz bellek: amnezik sendromun psikonomik bir çalışması, 1965, Academic Press, New York.
Thompson RF, Berger TW, Madden, IV, J. Memeli CNS'de hücresel öğrenme ve hafıza süreçleri, Ann. Rev. Neurosci., 1983, 6, 447-491.
Vinogradoua OS Bilgi ve limbik sistem kaydı. İçinde: Nöral aktivite ve davranışta kısa vadeli değişiklikler (eds. G. Horn ve R.A. Hinde), 1970, s. 95-140, Cambridge University Press, Cambridge.
Warrington E.K. Organik amnezide yetersiz tanıma belleği, Cortex, 1974, 10, 289-291.
Weiskrantz L., Warrington EK Amnezik hastalarda şartlandırma, Neuropsychology, 1979, 17. 187-194.
Zangwill OL Amnezik sendrom. In: Amnesia (editörler C. W. M. Whitty ve O. L. Zangwill), 1966, Butterworth, Londra.
Zangwill OL Hafıza bozuklukları. İçinde: Psikiyatri El Kitabı: 1. Genel psikopatoloji (eds. M. Shepherd ve OL Zangwill), 1983, s. 97-113. Cambridge University Press, Cambridge.
Zola-Morgan S., Squire L. R. Maymunlarda medial temporaMesions ile korunmuş öğrenme: motor ve bilişsel becerilerin korunması, J. Neurosci., 1984, 4, 1072—
Zolman JF Isıl güçlendirme ile genç çocuklarda ayrımcılığın öğrenilmesi, Psychol. Rec., 1968, 18, 303-309.
Zolman JF Uyaran tercihleri ve genç civcivlerde öğrenme ayrımcılığı biçimi, Psychol. Rec., 1969, 19, 407-416.
BÖLÜM 7
ÇOK FAZLA İZ DEPOLAMA SİSTEMİ! BEYİN ASİMETRİSİ VE İZLEME
Beynin sağ ve sol yarıkürelerinin basit yapay nesnelere damgalama sürecinde farklı roller oynayabileceğine dair ilk ipucu, PMIG'nin elektron mikroskobik çalışması sırasında elde edildi. Bu çalışma Sec.'de ayrıntılı olarak ele alınacaktır . 10.2. Kısacası, öğrenmenin sol PMVG'deki sinapsların yapısındaki değişikliklerle ilişkili olduğu bulundu, ancak sağ PMVG'de istatistiksel olarak anlamlı bir değişiklik gözlenmedi. Damgalama sırasında hemisferik asimetrinin bu kanıtı, beklenmedik olmasına rağmen , önceki bölümlerde açıklanan diğer damgalama deneylerinin sonuçlarıyla çelişmedi . İlk biyokimyasal çalışmalar (Bölüm 2 ve 3), hem sol hem de sağ PMVG'yi içeren beyin örneklerindeki biyokimyasal kaymaları analiz etti; radyootografik çalışma için, her iki alan da taraf dikkate alınmadan alındı (bölüm 4) ve bölüm'de açıklanan yıkım deneylerinde. 5, PMVG'ye her iki yarım kürede hasar verildi. Bu nedenle, tüm bu çalışmalarda aktif bölgenin sol DMAH olması ve sağ DMAH'nin yakalanmasının deneylerin sonuçlarını önemli ölçüde etkilememesi veya yalnızca mevcut değişikliklerin şiddetini azaltması mümkündür. Bu nedenle, tercih edilen uyaranın hafıza izlerinin korunmasında sağ ve sol PMVP'nin göreceli rolünü inceleyerek interhemisferik asimetriye ilişkin morfolojik verileri test etmek için özel bir dizi deney düzenlendi.
Sıralı hasar ile deneyler
Sol ve sağ LMWH'ler (lLMWH'ler ve pLMWH'ler) aslında damgalama sürecinde farklı işlevler yerine getiriyorsa, seçici yıkımlarının damgalama üzerinde farklı etkileri olmalıdır. Örneğin, bir tavuğun bir nesneyi tanıması sırasında belleğin temeli, lPMG'deki sinaptik değişiklikler ise, o zaman bu bölgenin yok edilmesi , bellek izlerinin korunmasını bozmalıdır. Aynı gerekçelerle, PPMVG bilgilerin saklanmasına katılmadığı takdirde, zararı yansıtılmamalıdır.
Resim: 7.1. Bu modelde sadece sol PMVG'nin iz depolama işlevine sahip olduğu ve civcivlerin ameliyattan önce eğitildiği varsayılmaktadır. İz depolama üzerinde çeşitli hasar etkileri tahmin edilmektedir. Kareler sağ ve sol PMVG'yi temsil eder. A. İlk işlem (/) sağ MRH'yi (rLRH) kaldırır ve civciv iyileştikten sonra tercih tepkisi ölçülür. Daha sonra ikinci operasyonda (2) sol DMAH (LMWH) çıkarılır ve ikinci testte tercih yanıtı ölçülür. B. Bu deneyde, önce lPMCH (/) ve ardından pPMCH (2) çıkarılır.
izleri kaydetme konusunda toplanın. Sadeliğiyle çekici olan ancak potansiyel olarak aşırı basitleştirme tehlikesi taşıyan bu araştırma hattı, sonraki bir dizi deneyde geliştirilmiştir (Cipolla-Neto ve diğerleri, 1982).
işlevine sahip olduğu ancak rLMVP'nin olmadığı hipotezini kabul edersek , çeşitli cerrahi müdahalelerin sonuçlarını tahmin edebiliriz (Şekil 7.1). Örneğin eğitimli tavuklarda PMSH'nin yok olması durumunda, eğitim sırasında kendilerine sunulan nesneyi tanımaları gerekir (Şekil 7.1, A, /). lPMHG'nin çıkarılmasıyla, bu tavuklar nesne için gelişmiş tercihlerini kaybetmeli ve hafıza kaybı göstermelidir (Şekil 7.1, A, 2). Şek. 7.1, B , önce lPMVG'nin ve ardından pPMVG'nin yok edilmesinin beklenen sonuçlarını gösterir. Bu çeşitli tahminler, Şekil 2'de gösterilen deneyde test edildi. 7.2. Eğitim prosedürü, önceki beyin yıkımı deneylerindekiyle aynıydı (bölüm 5.1 ve 5.2). Doğumdan itibaren karanlıkta tutulan civcivler , yaklaşık 19 saatlikken tepeden aydınlatmaya maruz bırakıldı. Daha sonra dönen kırmızı kutu veya doldurulmuş tavuk verilerek 2 saat boyunca eğitildiler. Eğitimden yaklaşık 3 saat sonra civcivler uyuşturuldu ve gruplara ayrıldı. Bazı
Test 1 Test 2
45 50 68
1 ■■ 1
5 i P -iI EZD -I 1 - i. i
Resim: 7.2. Ardışık hasar ile deneyin şeması. A. İlk olarak rPMCH ve ardından lPMCH ile yaralanan civcivleri test etme prosedürü . Beyin hasarı olan tüm yavru grupları için zaman aralıkları aynıdır. B. Sahte çalıştırılan kontrol grubu için prosedür. Gösterimler için Şekil 5L'deki başlığa bakın. (Cipolla-Neto ve diğerleri, 1982.) lPMCH'ye zarar verirken diğerleri rPMCH'ye zarar verdi. Grupların geri kalanı kontrol olarak görev yaptı. Bunlardan birinde Wulst sol yarımkürede, diğerinde - sağda hasar gördü. Sahte ameliyat edilen grup, daha önce açıklandığı gibi hazırlandı. Tavuklar ameliyattan çıktıktan sonra seri tercih testi (Test 1) uygulandı ve tercih puanı hesaplandı (s. 104). Tavuklar daha sonra tekrar uyuşturuldu ve kalan simetrik yapının (PMWG veya Wulst) zarar gördüğü ikinci bir ameliyat yapıldı . Böylece hasar iki taraflı hale geldi. Bundan sonra civcivler karanlık inkübatöre geri konuldu ve anestezinin etkisi geçince ikinci bir tercih testi uygulandı (test 2). Tavuklar anestezi altında kalırken kodlandı; "kör yöntemin" davranışını daha fazla inceledi.
kırmızı kutu ve doldurulmuş tavukla sunulan tavuklarda tercih puanları arasında anlamlı bir fark göstermedi . Bu nedenle, bu göstergeler ilgili gruplara göre birleştirilmiştir. İlk önce sol veya sağ Wulst'un yok edildiği civciv grupları arasında da önemli bir fark yoktu , bu nedenle bu iki grup için veriler de birleştirildi.
Tercih testinin sonuçları Şek. 7.3. Sahte ameliyat edilen tavuklar, her iki testte de tanıdık uyaranı tercih ettiler. Test 2'deki ortalama tercih puanları, test 1'dekinden biraz ama yine de önemli ölçüde düşüktü. Wulst'tan zarar görmüş civcivler, test 1 ve 2'deki kontrollere benzerdi. Bu lezyona sahip kuşlar, her iki testte de tanıdık nesneyi tercih etti. Ancak, kontrol grubunun aksine, Wulst'u hasar görmüş kanatlıların ortalama puanları test 1 ve 2'de sabit kaldı.
Hipoteze göre, tek taraflı pPMCH hasarı olan civcivler, test 1'deki tanıdık uyaranı tercih etmiş olmalıdır (Şekil 7.1, A,/). Bu varsayım doğrulandı (Şekil 7.3, L, pPMVG). Ortalama tercihleri, sahte ameliyat edilen grup veya tek taraflı Wulst lezyonları olan gruptan önemli ölçüde farklı değildi. Kalan lPMCH'nin çıkarıldığı ikinci işlemin beklenen sonucu Şekil 1'de gösterilmiştir. 7.1, K, 2. Bu tavuklarda aslında gözlenen tanıdık nesne tercihi ortadan kalkmış olmalıdır . Test 2'deki ortalama tercih puanları, rastgele 50 seviyesinden önemli ölçüde farklı değildi (Şekil 7.3b , pPMCG). Bu testteki performansları da sahte olarak çalıştırılan kontrol grubundan ve Wulst'u zarar görmüş tavuklardan önemli ölçüde daha düşüktü. Dolayısıyla, bu sonuçlar RCS'de tasvir edilen modelle tutarlıdır.
100 gram
Bir Test 1

0 <U. 1— G/zzzh
lPMWH LPMWH W
/V: 52 P 17 16
Resim: 7.3. Farklı civciv grupları için ortalama tercih puanları. Veri**, sol ve sağ Wulst lezyonları (W) birleştirilmiş civcivler için. Sağ veya sol PMVH'nin ilk önce yok edildiği civcivler sırasıyla rMVVG ve lMVVG olarak adlandırılır. Test 1'deki gruplar arasında ortalama puanlarda anlamlı bir fark yoktu. Test 2'de, pPMCH grubunun ortalama tercih puanları, 1) sahte ameliyat grubu (P<0.05), 2) Wulst hasarı olan tavuklar ( P<0.005) ve 3) lPMCH grubununkinden önemli ölçüde daha düşüktü. (P<0.05). ). Test 2'deki pPMVP grubu için ortalama tercih puanları, 50'den önemli ölçüde farklı değildi. Diğer tüm tavuk grupları için ortalama puanlar, bu şans seviyesinden önemli ölçüde yüksekti ve birbirinden önemli ölçüde farklı değildi. (Cipolla-Neto ve diğerleri, 1982.)
yani, LMVP'nin bir depolama işlevi gerçekleştirirken RPMV'nin gerçekleştirmemesi.
Bu modelin en basit varyantında, yalnızca LPMG bir nesneyi tanımak için gereken bellek izlerini saklar. Bu doğruysa, civcivler eğitimden sonra lPMG'nin çıkarılması üzerine hafıza kaybı geliştirmelidir (Şekil 7.1, B, /). Deneyde bu sonuç elde edilememiştir. Hasarlı ILMCH'li civcivler, eğitimden kısa bir süre sonra tanıdık nesneler için belirgin bir tercih gösterdiler (Şekil 7.3, A, lLMCH). Bu nedenle tanıma için gereken bilgiler yalnızca lPMG'de depolanamaz. Bölümde verilen morfolojik verilere rağmen mümkündür . 10.2, tanıma her iki PMVP tarafından sağlanır ve her biri eğitim sırasında geliştirilen tercihten sorumludur. En az bir PMVG sağlam kaldığı sürece civciv tanıdık nesneyi tercih etmeye devam eder. Buradan, bu PMVG'nin çıkarılmasının amneziye neden olması gerektiği sonucu çıkar. Önce pPMCH, ardından lPMCH hasar gördüğünde olan tam olarak budur (Şekil 7.3, pPMCH). Önce lPMCH'yi ve ardından pPMCH'yi kaldırırsanız ne olur? Tanıdık nesne tercihi devam etti (Şekil 7.3, B, lPMCH). Hasarlı lPMCH'ye sahip kuşlar için ortalama değerler, sahte olarak çalıştırılan kuşlar ve hasarlı Wulst'a sahip grup için olanlardan önemli ölçüde farklı değildi . Ayrıca, test 2'de, lPMCH grubu için ortalama puanlar (ilk olarak IPMCH'nin hasar gördüğü), pLMCH grubu için olanlardan önemli ölçüde daha yüksekti. Bu sonuçlar , sağ ve sol SHG'lerin bilgileri aynı ölçüde depoladığı varsayımını desteklememektedir. Aslında, izlerin yalnızca LRHG'de depolanması ve her iki LRH üzerinden dağıtılan depolama olmak üzere iki modelin hiçbiri elde edilen sonuçları açıklayamaz.
Açıklamanın karmaşıklığı, test 2'de her iki grubun civcivlerinin iki taraflı PMVG tahribatına tabi tutulmuş olması gerçeğiyle özellikle vurgulanmaktadır. Bununla birlikte, bir grupta (lPMCH) tanıdık uyaran için belirgin bir tercih korunurken , diğer grupta (rPMCH) tepki rastgeleydi. Tanıdık bir nesneyi tercih eden tavuklarda tanımanın korunduğunu varsayarsak, sonuç olarak, DMAH grubundaki tavuklarda, bellek her iki DMAH'ın yokluğunda da çalışabilir.
Bu durumda ne olduğu iki gerçek temelinde açıklanabilir. İlk olarak, deneylerin son aşamasında PMVP'de aynı iki taraflı hasara sahip olan tavuklarda, bu durum farklı şekillerde elde edildi. Bir grupta önce lPMCH, ikinci grupta rPMCH hasar gördü. İkincisi, öğrenmeden önce PMVP'ye verilen iki taraflı hasarın bir tercih geliştirmeyi imkansız hale getirdiği daha önce gösterilmişti (Bölüm 5.1). Bu civcivlerde, tercih puanları rastgele 50 seviyesine yakındı (Şekil 5.3). Antrenman sonrası bilateral yaralanma yapıldığında ortalama tercih skoru 64.6 ± 7.53 idi. Bu gösterge, 50'den önemli ölçüde farklı değildir (Z=l.91, t — Student testi). Rastgele böyle bir değer elde etme olasılığı t 1/ 20'den fazla ama 1/10'dan az . Başka bir deyişle, fark gerçeğe yakındı ve bu, tanıdık nesne hakkında bazı bilgilerin saklandığını gösteriyordu. SMCH'nin bilgilerin kaydedilmesine ve saklanmasına dahil olması, ancak öğrendikten bir süre sonra bilgilerin yavaş yavaş onu terk etmesi mümkün mü? Başka bir deyişle ? .
Zaman
Antrenman sırasında ve hemen sonrasında
Mezun olduktan yaklaşık 3 saat sonra
Mezuniyetten yaklaşık 26 saat sonra
Resim: 7.4. Tercih izlerinin korunmasında sol ve sağ PMVP'nin rolünün modeli . Bir nesneyi tanımak için gerekli bilgilerin her iki SHG'de de saklandığı varsayılır. Bu durumda, lPMSH , en azından metinde açıklanan deney süresince kalıcı bir depolama (X) olarak kabul edilir ve rTMVG, bilgileri yavaşça başka bir depoya aktaran bir tampon, geçici depolama (BX) olarak kabul edilir. , X'. Eğitimin bitimini takip eden 3 saat içinde nispeten az bilgi aktarılır; tam transfer eğitim bitiminden 26 saat sonra tamamlanır.
Yukarıda önerilen modellerin hiçbirinde hesaba katılmayan , iz depolamanın dinamik bir unsuru var mı ?
Hem yukarıda açıklanan beyin yaralanmalarıyla ilgili deneylerin sonuçlarına hem de ardışık yaralanmalarla yapılan deneylere karşılık gelen bir model, Şekil 1'de gösterilmektedir. 7.4. Bu modele göre, eğitim sırasında bilgi her iki TMGS'de de saklanır. Öğrenmeden önce onları yok etmek , tercihlerin oluşamaması için izleme deposunu ortadan kaldırmalıdır. lPMG'de bilgilerin depolanması, en azından deney süresi boyunca "kalıcı" kabul edilir. PPMG'deki depolama , bilgileri başka bir X' deposuna ileten "arabellek" olarak kabul edilir. Öğrenmeden sonra her iki PMSH de kaldırılırsa, izlerin gücü ne kadar bilginin X' deposuna çevrildiğine bağlı olacaktır: aralık ne kadar uzunsa, tanıdık olana yönelik tercih o kadar güçlü olacaktır. Deneysel veriler, aralık yaklaşık 3 saat olduğunda, aktarılan bilgi miktarının tanıdık bir nesne tercihini sürdürmek için yeterli olmadığını göstermektedir (Şekil 5.6, PMVP). Bu nedenle, eğitimden yaklaşık 3 saat sonra bir PMIG çıkarılırsa izlerin tamamen korunması beklenebilir. Modele göre (Şekil 7.4), eğitimden 3 saat sonra rPMG silindiğinde , ara depolama imha edilecek ve artık bilgi X'e iletilmeyecektir. Bu durumda, nesne tanıma esas olarak depolamaya bağlı olacaktır.
Test 1 Test 2
19 20 24 45 50 68
A ben P—i ben • ■■ 1
B I R— ben ben 1 !=□ 1
Resim: 7.5. Tavukların eğitildiği deney şeması karanlık inkübatöre geri döndürüldü ve ardından PMVP iki taraflı olarak yok edildi. A. PMVG'si zarar görmüş civcivler. Kontrol grubu. (Cipolla-Neto ve diğerleri, 1982.)
lPMVG. İkinci operasyonda bu bölge kaldırılırsa, X' içine ne kadar bilgi gittiğine bağlı olarak ortalama tercih puanı aşağı yukarı 50'ye yakın olacaktır. Ortaya çıkan ortalama puan (Şekil 7.3, B, pPMCG), 50'den önemli ölçüde yüksek değildi; bu, eğitimden 3 saat sonra varsayılan X' deposuna nispeten az bilginin aktarıldığını gösterir. Eğitimden 3 saat sonra yalnızca lPMCH yok edilirse, rPMCH'deki ara depolama bozulmadan kalır ve bilgi X' içine akmaya devam edebilir. Bu aktarım tamamlandı mı, tamamlandıysa ne zaman tamamlandı? pPMCG'deki ikinci yıkımın, eğitimin bitiminden yaklaşık 26 saat sonra gerçekleştirilmesi durumunda (Şekil 7.2), tanıdık bir nesne için ortalama tercih değerleri, kontrol grubununkinden neredeyse ayırt edilemez (Şekil 7.3). , B, lPMCH). Bu sonuç, antrenman bitiminden 26 saat sonra transferin tamamlandığını düşünmemizi sağlar.
Şek. 7.4, varsayılan X' bilgi transferinin tamamlanmasının ardından, PMVP'nin iki yönlü olarak kaldırılmasının tercih izlerinin korunmasını etkilememesi gerektiği varsayılmaktadır. Bu tahmin , şeması Şekil 1'de gösterilen deneyde doğrulandı . 7.5. Tavuklar , önceki deney serilerinde olduğu gibi yetiştirildi ve eğitildi. Eğitimin tamamlanmasından yaklaşık 21 saat sonra civcivlere bir tercih testi (Şekil 7.5, testi) verildi, ardından karanlık inkübatöre geri gönderildi ve eğitimden 26 saat sonra anestezi uygulandı. Sahte ameliyat edilen kontrol grubu 30 civcivden oluşuyordu. 30 deneysel tavukta PMVG'nin iki taraflı imhası gerçekleştirildi. Civcivler ameliyattan sonra karanlık bir inkübatöre yerleştirildi ve 68 saatlik olduklarında ikinci bir tercih testine tabi tutuldular (Şekil 7.5, test 2).
Tercih göstergeleri, şek. 7.6. Tavuklara anestezi uygulanmadan önce gerçekleştirilen test 1'deki iki tavuk grubu için ortalama değerler önemli ölçüde farklılık göstermedi. Test 2'de, her iki grup için ortalama tercih puanları, test 1'den önemli ölçüde daha düşüktü. Deney grubunun ortalama puanı, sahte grubunkinden biraz daha düşük olmasına rağmen, bu fark istatistiksel olarak
Resim: 7.6. Ortalama tercihler. Test 2'de, her iki grubun ortalama tercih puanları test 1'dekilerden anlamlı derecede düşüktü (P<<0.05). (Cipolla-Neto ve diğerleri, 1982.)
güvenilmez. Model tarafından tahmin edildiği gibi, deneysel tavuklar, tanıdık uyaran için önemli ölçüde yüksek düzeyde bir tercih gösterdiler (P<0.001).
Önerilen model , deneysel verilerin çoğunu oldukça iyi açıklıyor ve tahminlerinden biri doğrulandı. Ancak bunun olası modellerden sadece biri olduğu vurgulanmalıdır. lPMSH'nin rolüne ilişkin varsayımlar, deneylerle doğrulanmıştır (Bölüm 4 ve 5). Bununla birlikte, eğitimden 26 saat sonra PMCH'nin iki taraflı olarak çıkarılması veya LMCH ve rPMH'ye sıralı hasar verilmesi izlerin korunmasını tehlikeye atmadığı için, X'in varlığı yalnızca davranışsal verilere dayanarak önerilir . Bu deneysel verilerden, SMVG'nin dışında bir iz depolama sistemi olması gerektiği sonucu çıkar. Bu sistemde depolanan bilgiler, pPMCH eğitimden kısa bir süre sonra yok edilirse eksik kalır (Şekil 7.3b, pPMCH). Bu sonuçlar, izlerin X' içinde depolanmasının, yalnızca sPMHG korunursa oluşturulabileceğini ve bunun bir " tampon" depolama işlevi olduğu fikrinin dayandığını göstermektedir.
Birçok bakımdan bu model eksiktir. Örneğin , böyle bir konumun SPGM olmaması dışında, X' konumu hakkında hiçbir deneysel veri yoktur . SMVP'den bağlantıların yansıtıldığı alanlarda X' aramak makul olacaktır . Ayrıca, normal koşullar altında rPMG'nin kalıcı bir depolama işlevi görmediği varsayılır, çünkü imprinting sırasında sinapslarda istatistiksel olarak anlamlı bir değişiklik bulunmamıştır (Bölüm 10.2). Ancak, bu olumsuz kanıt yeterince güvenilir değildir ve bu tür değişiklikler daha sonraki çalışmalarda bulunabilir. Bunların yokluğunda, PMWG'nin bilgileri diğer depolama sistemlerine aktaran geçici bir depolama olduğu varsayılmalıdır.
Bellek izlerini geçici depolardan kalıcı olanlara aktarma fikri bilgisayarlarda yaygın olarak kullanılmaktadır ve bu açıdan oldukça önemsizdir. Sinir sistemini inceleyen bilimler için de yeni değil . Örneğin, Marr (Mugg, 1970, 1971), memelilerdeki neokorteks ve arkikorteksin (hipokampus dahil) bellek süreçlerinde işlev ayrımı gerçekleştirdiğini öne sürdü. Bu yazar, arki-korteksin mevcut bilgilerin geçici bir deposu olduğuna ve kalıcı izlerin neo-kortekste depolandığına inanıyordu; Archicortex'ten neokortekse bilgi aktarımı uyku sırasında gerçekleşir. Bundan • izlerin orijinal deposunun onları en az bir gün tutabilmesi gerektiği sonucu çıkar. Marr'ın modeli spekülatif kalsa da , bilginin kalıcı bir depolama sistemine aktarıldığı geçici bir depolama kavramı sinirbilimde oldukça yaygındır .
Bilginin X'e aktarılması için gereken süre nispeten uzundur ve bu işlemin gerçekleştirildiği fizyolojik süreçler bilinmemektedir. Daha yakın zamanlarda, bu tür süreçler tamamen gizemliydi. Şimdi çok dikkat çekiyorlar ve fizyolojik analiz için erişilebilir durumdalar. Örneğin, çok sayıda nöron, merkezi sinir sisteminde aracılar olarak görev yapan nöropeptitler içerir. Nöropeptitler , sinapsta geri dönüştürülmemeleri veya geri yüklenmemeleri bakımından asetilkolin gibi bir dizi diğer aracıdan farklıdır . Nöropeptidler hücre gövdesinde sentezlenir ve akson boyunca taşınır. pPMVC'de bulunan peptiderjik nöronların , afferent lifler tarafından uyarılması mümkündür. Bu giriş sinyali , verilen peptidi kodlayan haberci RNA'nın sentezini indükleyebilir veya hızlandırabilir . Daha sonra akson boyunca taşınacak ve terminallerden salınacak, postsinaptik nörondaki reseptörlere bağlanacak ve diğer nöronlardaki reseptörlere difüzyonla yayılacaktır. Reseptör aktivasyonu, postsinaptik nöronlarda metabolik kaymalara neden olabilir ve bu, örneğin, subsinaptik zar üzerindeki reseptörlerin yoğunluğunda bir değişikliğe yol açabilir. Tabii ki, bunların hepsi saf spekülasyon. Ancak bahsi geçen fizyolojik süreçlerin birçoğu merkezi (Potter ve diğerleri, 1981; Chen ve diğerleri, 1983) ve otonomik (Zigmond ve Chalazonitis, 1979) sinir sistemlerinde tanımlanmıştır. sPMSH'de afferent impulsların ortaya çıkmasından, peptit sentezindeki değişikliklere, bunun akson boyunca taşınmasına , salınmasına ve postsinaptik hücre üzerindeki etkisine kadar varsayılan olay dizisinin tamamlanması saatler alabilir. Bu nedenle, farazi X' içindeki izlerin yavaş oluşumu oldukça anlaşılırdır.
Hemisferik fonksiyon modifikasyonu
Seri yaralanma serilerinde (Şekil 7.2) kullanılan tüm cerrahi müdahaleler önceden eğitilmiş tavuklar üzerinde gerçekleştirilmiştir. Eğitimden önce PMSH'ye ilk hasar verildiğinde farklı sonuçlar elde edildi . Civcivlerin eğitim süresi boyunca en az bir PMVP'yi koruyarak nesne için belirgin bir tercih geliştirdiğini hatırlayın (Bölüm 5.3) . Aynı zamanda, SMVG'nin hangi tarafta tutulduğu * önemli değildi. Bu , Şekil 1'de gösterilen modele göre beklenmelidir. 7.4: lPMGP korunursa, depolama sağlayabilir ve pPMGP korunursa, pPMGP ve X'in birleşik eylemiyle aynı şey elde edilebilir. Bölümde açıklanan deneylerde. Şekil 5.3'te, bir PMVG'nin imha edilmesinden sonra eğitilen civcivler, ardından 1'e yeniden anestezi uygulandı ve ikinci PMVG hasar gördü (Şekil 5.10). Modele göre (Şekil 7.4), LRHG izlerin deposudur ; eğitim sırasında tek başına sağlam kalırsa , sonraki yıkımı amneziye yol açmalıdır . Bu varsayım doğrulandı (Şekil 7.7, B, L). Bu nedenle, pLMCH eğitimden önce (Hörn ve ark., 1983) veya eğitimden kısa bir süre sonra (Cipolla-Neto ve ark., 1982) çıkarılırsa, izlerin korunması kritik olarak ILMCH'nin korunmasına bağlıdır. Bu* veriler modelle tutarlıdır. Bu model, civcivlerin bir pPMVP'yi korurken eğitilip eğitilmediğini ve ardından; bu yapı zarar görmüş mü? Eğitimin bitiminden hasar anına kadar yeterince uzun bir süre geçmişse , X'te izlerin oluşumu tamamlanmalıdır, bu da tercih tepkisinin korunmasını sağlayacaktır. Eğitimin bitiminden yaklaşık 20 saat sonra bu tür bir hasarın sonuçları Şekil 1'de gösterilmektedir. 7.7 B. Bu Civcivler için ortalama değerler rastgele seviyeden önemli ölçüde farklı değildi. Bu nedenle deney, * X' içindeki izlerin oluştuğuna inanmak için hiçbir sebep vermiyor.
X' içinde izlerin oluşamamasının olası bir nedeni , rPMVG'nin çok erken yok edilmesi olabilir. Eğitimin bitiminden sonra yaklaşık 26 saat boyunca rTMG bozulmadan kalırsa varsayılan iz deposu bir tercihi koruyabilir (Şekil 7.3b, lTMG). X' de izlerin oluşması için belki 20 saat yeterli değildir? Eğitimden 3 saat sonra pPMCH, lPMCH ile birlikte çıkarıldığında.

Resim: 7.7. PMVG'nin tek taraflı olarak yok edilmesinden sonra eğitilen civcivlerde ortalama tercih puanları . Bu deneyin şeması, Şek. 5.10. incirden farklı olarak 5.11 ortalama tercih puanları, eğitim ve Test 1 sırasında lPMCH (L) veya rPMCH (P) sağlayan civcivler için ayrı ayrı gösterilir. Test, eğitimin tamamlanmasından kısa bir süre sonra gerçekleştirildi . 50'den ortalama puanın kritik farkı (P^0.05), B'de sahte ameliyat edilen tavuklar için puanın sağında gösterilir. Sahte ameliyatlı piliçlerin ortalama puanlarını 50 ile karşılaştırmak için kritik fark (P^0.05) karşılık gelen çubuğun ( LO) sağında B'de gösterilir . lPMCH ve pPMCH gruplarını kontrol grubuyla karşılaştırmak için kritik fark (P=0.05), her grafiğin sağındaki dikey parantez ile gösterilir. A. Test 1. Tüm ortalama tercih puanları anlamlı olarak 50'den büyüktür (P < 0.005) ve birbirinden farklı değildir. B. Test 2'deki Ortalama Tercih Puanları, civciv kalan PMA'yı çıkarmak için yapılan ameliyattan iyileşirken gerçekleştirilir. Sahte ameliyat edilen tavuklar tanıdık nesneyi tercih ettiler. Beyin hasarı olan tavuklarda nesne tercihi yoktu ve ortalama değerleri rastgele seviyeden önemli ölçüde farklı değildi. Etkilenen kuşların iki grubu için ortalama tercih puanları , kontrol grubundan önemli ölçüde farklıydı. (Horn ve diğerleri, 1983.)
tercih indeksi 64.4 ± 7.53 idi (Şekil 5.6, PMVG). 20 saat boyunca bir pPMCH'nin varlığı, X'e bir miktar bilgi aktarımı sağlıyorsa, o zaman pPMCH'nin çıkarılmasından sonraki ortalama tercih 64'ten yüksek olmalıdır. Aslında, 59.39 ± 9.73 idi (Şekil 7.6, B, P ) . Bu nedenle, veriler X' içinde izlerin oluşumunu desteklemez. Bu sonuç, X'e transferin öğrenmenin bitiminden 6 saat sonra tamamlandığını gösteren yeni bir çalışmayla (Davey, McCabe, Horn, 1987) desteklenmektedir . Özetlemek gerekirse (Şekil 7.8, B): yalnızca pPMVP'nin korunmasıyla, normal koşullar altında bir depo işlevini yerine getirerek izleri korumayı öğrenmede kritik hale gelir.
mevcut sadece lPMWG; X' içinde PMSH'den bağımsız olarak işlev gören hiçbir iz oluşumu yoktur .
Evcil tavuklarda gözlemlediğimiz sağ yarımküre fonksiyonlarındaki değişiklik diğer türlerde de mevcuttur. Nottebohm ve diğerleri ( 1976 ), ventral hyperstriatum'un (CVH) kaudal kısmının kanaryada (Serinus canarius) şarkı söylemenin düzenlenmesinde yer aldığını bulmuşlardır. Bu bölge , bazı ötücü kuşların beyinlerinde bulunan özelleşmiş bir çekirdektir, ancak evcil tavukların ventral hiperstriatumunda karşılığı yoktur. Nottebohm ve diğerleri (Nottebohm ve diğerleri, 1976; Nottebohm, 1980) , sol CVG'nin şarkı söyleme öğreniminde baskın bir rol oynadığını ileri sürmüştür. Sol CVG'nin yok edilmesinden sonra , sağ CVG ve onun efferent yolları bu işlevi üstlendi ve bunun sonucunda kuşlarda normal ötüş geri geldi. Böylece, sol CVG'yi yok ederek, hemisferik hakimiyetin yeniden düzenlenmesini sağlamanın mümkün olduğu ortaya çıktı (Nottebohm, 1980). Benzer sonuçlar, konuşma işlevi için genellikle sol yarıkürenin baskın olduğu insanlarda da gözlemlenmiştir . Bir yetişkinde bu yarımküre çıkarılırsa veya buna karşılık gelen bölgeler yok edilirse, böyle bir hastada ciddi bir konuşma bozukluğu olur ve telafi olasılığı genellikle minimumdur (bkz . Moscovitch tarafından yapılan inceleme, 1977). Buna karşılık çocuklarda sol yarıküre çıkarıldığında veya hasar gördüğünde konuşma işlevi sağ yarıküre tarafından yürütülebilir (Dennis ve Kohn, 1975; Hosen, 1976; Trevarthen, 1983 ). Böylece, sağ yarıküre normalde sol yarıküreye özgü işlevler kazanır, ancak bu yalnızca sol yarıküre yaşamın erken dönemlerinde hasar görürse gerçekleşir.
Civcivlerde, öğrenmeden önce lPMSH yok edilirse sağ hemisfer işlevinde bir miktar azalma olduğu da görülmektedir ; X' içindeki izleme depolaması yapılandırılmamış. Benzer bir fenomen , insan beyninde de gözlemlenir, sağ yarıküre, sol yarıkürenin erken hasar görmesi nedeniyle, kendisi için alışılmadık işlevleri yerine getirmek zorunda kaldığında. Yetişkin bir insanın sağ yarıküresi, sol yarıküreden daha gelişmiş görsel-uzaysal işlevlere sahiptir (bkz . Levy tarafından yapılan inceleme, 1974). Sağ hemisferin bu işlevleri , konuşma işlevlerinin yerine getirilmesini üstlenirse azalır (Levy, 1969).
/. sinir modeli
Sol yarıkürenin çıkarılmasının sağ yarıkürenin işlevini değiştirdiğine dair kanıtlar , beynin iki yarısı arasında bir etkileşim olduğunu düşündürür. Hücresel me hakkında- 11-140


AB _
Resim: 7.8. Tavuk beyni diyagramı, üstten görünüm. Her yarım küredeki oval kontur, PMVP'yi belirtir. Öğrenme sırasında oluşan bir bellek sistemi modeli (E) ve hasar durumunda değiştirilmeleri (B) sunulmaktadır. A. Depolama sistemlerinden (X) birinin kritik halkası lPMHG'dir. Bilgiler, geçici bir depolama (TS) görevi gören rTMVG'nin etkisi altında X'e aktarılır . Ayrıca bkz. 7.4. B. Antrenmandan önce lPMSH'yi (gölgeli alan) kaldırmanın sonucu. İkinci depolama sistemine (X') aktarım gerçekleşmez ve rPMG, izlerin korunması için kritik hale gelir. P - sağ taraf; L - sol taraf. (Horn ve diğerleri, 1983.)
Bu etkileşimin mekanizmaları hakkında neredeyse hiçbir şey bilinmiyor. Bununla birlikte, sonraki deneyleri kurmak için verimli olabilecek olası mekanizmaları hayal etmek zor değil . Örneğin, Şekil l'de gösterilen sistemi ele alalım. 7.9. Hücre gövdeleri N, P, L ve Q PPMVG'de bulunmaktadır. Böyle bir sistem sağlam bir civcivde nasıl çalışır (Şekil 7.8, K), pPMCH'nin bilgileri X'e ileten geçici bir depolama görevi görmesi beklenirken ve ayrıca öğrenmeden önce lPMCH çıkarıldığında (Şekil 7.8, K). B), pPMCH kalıcı bir depolama gerçekleştirdiğinde ve X' içindeki izler oluşmadığında? Şek. 7.9, uyaranın sinir izinin oluşum mekanizması dikkate alınmaz (bölüm 10.3). Bir uyaranın, uyaranın etkisinden sonra devam eden özel bir nöron grubunda değişikliklere neden oluyorsa, sinir sisteminde iz bıraktığını varsaymak yeterlidir. Bu değişiklikler kısa süreli geri dönüşlü ise bu iz geçici, uzun süre stabil ise kalıcı olacaktır.
Şekil l'de gösterilen her hücre 7.9, bir hücre popülasyonunu temsil eder. Nöron P , X'teki hücrelere bağlıdır; sadece aynı anda N ve L hücreleri tarafından etkileniyorsa boşalır.Neuron N, PMCG'deki geçici depolamanın efferent hücresidir. Nöron L doğrudan veya dolaylı olarak lPMCH ile ilişkilidir. Bu hücre, P hücresinin deşarjlarını düzenler ve böylece
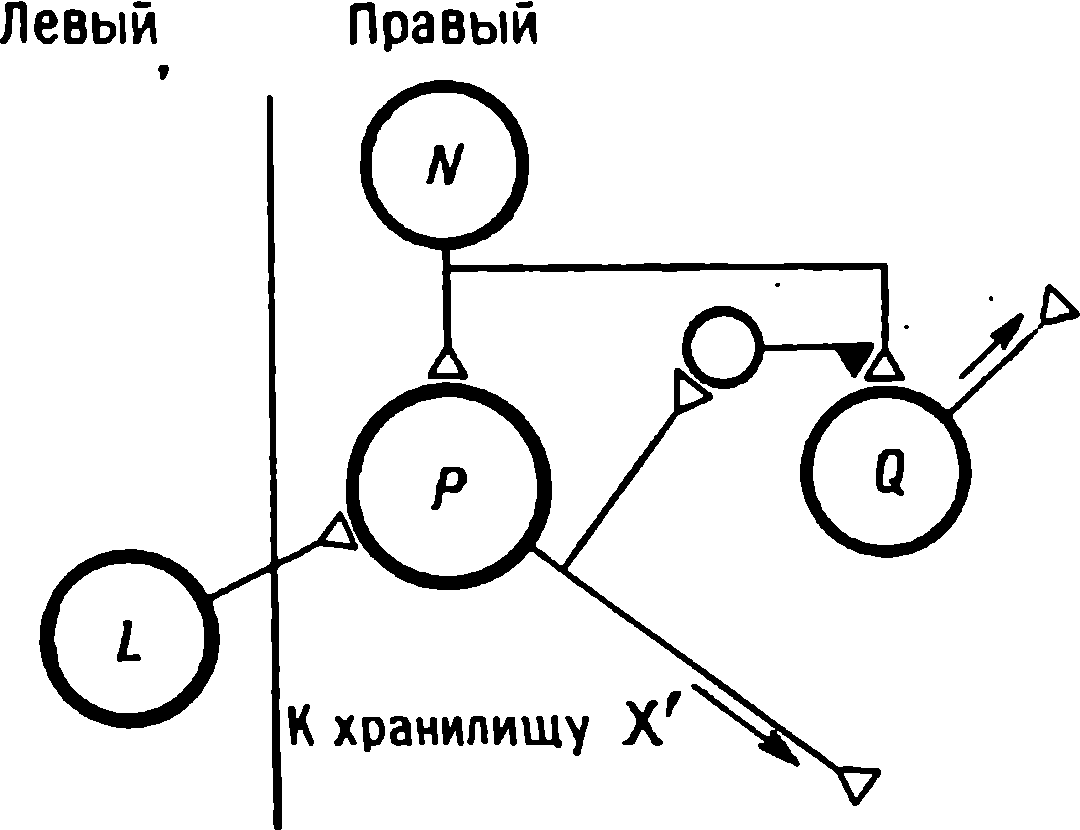
Resim: 7.9. Bellek izlerinin oluşumu sırasında lPMCH'nin rPMCH üzerindeki etkisini açıklayabilen olası sinirsel etkileşimlerin şeması . Beyaz üçgenler uyarıcı sinapslardır , siyah üçgen ise inhibe edici sinapslardır . Q hücre çıkışı sPMGB'deki diğer nöronlara veya beynin diğer bölümlerine, belki de yaklaşma-kaçınma davranışını düzenleyenlere gider. Daha ayrıntılı açıklamalar için metne bakın.
böylece belirli bilgilerin X' içindeki hücrelere transferini kontrol eder. Baskılayıcı bir uyarana maruz kalma süresi boyunca, N ve L hücreleri aynı anda uyarılır. P hücresi , uyaranın tüm süresi boyunca uyarıları boşaltır ve üretmeye devam eder . P hücresi yukarıda açıklanan özelliklere sahiptir (s. 158), yani uygun uyarma ile nöropeptidin sentezi için gerekli olan haberci RNA'nın sentezi artar. Bu nöropeptit, P hücresi aksonu boyunca taşınır ve sinaptik terminalinde salınır. Nöropeptit, X' nöronlarındaki reseptörlere bağlanır ve böylece kalıcı bellek izlerinin temeli olan plastik kaymaları indükler . Şek. Şekil 7-9'da, N hücresi aynı zamanda Q hücresini de ateşler P hücresi aksonunun teminatı yoluyla internöron I'i ateşler . Bu sinapsta iletimin hızlı ve "rutin" olduğu varsayılır, çünkü hücre I'de P hücresi tarafından salgılanan nöropeptit için reseptör yoktur . Hücre I , terminallerde N ve Q hücreleri arasındaki iletimi bloke eden bir inhibitör aracı salgılar. P hücresi boşaldığında , Q hücresi sessizdir. Şimdi lPMG'nin eğitimden önce çıkarıldığını varsayalım. Bu durumda, L hücresi işlevini durduracaktır . N hücresinin uyarımı eşiğin altında olduğu için P hücresi boşalmayacaktır . Sessizliği nedeniyle X'te hiçbir iz oluşmuyor . Aynı nedenle, N hücresinden gelen bir sinyal Q hücresini uyarır Q hücresinin bu deşarjının A' ve Q hücreleri arasındaki sinaptik temasta yapısal değişikliklere yol açtığı varsayılır (Bölüm 10.3.4). Bu değişiklikler, izlerin PMWP'de kalıcı olarak saklanmasının temelidir. yani 11*
LPMVG'nin yokluğunda bilgi PPMV*'de depolanır ve X'e aktarımı gerçekleşmez.
, Şekil 2'de gösterilen sonuçları sağlamalıdır . 7.8, B. Eğitimden sonra lPMSH'nin çıkarılması, X' izinin oluşumunu etkilemeyecektir, çünkü bu işlemi başlatmak için P hücresindeki gerekli değişiklikler eğitim süresi boyunca meydana gelmelidir . Şema ayrıca diğer beyin hasarı deneylerinin sonuçlarını da açıklayabilir. Eğitimden önce PMWG'nin iki taraflı olarak çıkarılması, Şekil 1'de gösterilen tüm sistemi ortadan kaldıracaktır. 7.9, böylece X' içindeki iz oluşturulamaz . Antrenmandan sonra rPMCH'nin çıkarılması (LPCH ile veya olmadan) çeşitli faktörlere bağlı sonuçlar doğuracaktır. Bu faktörler, nöropeptit sentezini artırmak için gereken süreyi , P hücresi aksonu boyunca taşınma hızını, taşınması gereken mesafeyi ve X' hedef hücresini etkilemek için gereken süreyi içerir. Örneğin, akson taşıma hızının 2 mm/saat (Thoenen ve Kreutzberg, 1981) olduğunu ve sağ yarım kürenin herhangi bir en dış noktası ile rPMSH'nin merkezi arasındaki maksimum mesafenin 6 mm olduğunu varsayalım. Yeni sentezlenen nöropeptitin en uzaktaki hedefe ulaşması için gereken süre 3 saat olacaktır.Taşınan nöropeptit ortaya çıkmadan önce P hücresinin terminallerinde bulunmadığını veya miktarının hücreyi etkilemeyecek kadar az olduğunu varsayarsak. X' hücreleri, daha sonra P hücresinin X' üzerindeki değiştirici etkisi, nöropeptit sentezinden 3 saat sonra başlayacaktır. rPMCG'nin hasar görmesi, P hücresini yok edecek ve aksonu boyunca taşımayı durduracak veya yavaşlatacaktır. Bu nedenle, eğitimin bitiminden 3 saat veya daha fazla sonra pPMWG hasar görürse, peptit zaten X'te görünecek ve bazı değişiklikler orada başlayacak. Hasar nöropeptit taşınması ve X'deki hücreler üzerindeki etkisi tamamlandıktan sonra yapılırsa, X'te izlerin oluşumu tamamlanana kadar devam edecektir. Birkaç saat sonra PMVP'nin ikili olarak çıkarılması , geliştirilen tercihin sürdürülmesine engel olmayacaktır . Bu düşüncelerin ışığında, nöropeptit metionin-enkefalini içeren hücre gövdelerinin PMWG'de bulunması ilginçtir. Bu hücrelerin bazılarının 5 mm uzunluğa kadar aksonları vardır (McCabe ve Hunt, 1983).
Yukarıda açıklanan şema çeşitli şekillerde test edilebilir . Örneğin, çalışması için gerekli bir koşul, sPMGP ile lPMGP arasında etkileşimlerini sağlayan bir bağlantının varlığıdır. Pek çok dolaylı ilişki olmasına rağmen aralarında doğrudan bir ilişki olduğuna dair bir kanıt yoktur (Bradley ve diğerleri, 1985). Bu bağlantılar işlevsel mi ve lPMCH, rPMCH'deki nöronları uyarabilir mi? Orada, yalnızca lPMCH ve pPMCH'den eş zamanlı bir girdi olduğunda ateşlenen P hücresi gibi nöronlar var mı? Belirli nöropeptitlerin seçici blokajı, antrenman sırasında ve tamamlandıktan sonra 6 saat veya daha uzun bir süre boyunca her iki PMIG korunsa bile X'te eser oluşumunu bloke edecek mi? Elbette şema yanlış olabilir. Ancak şu anda, nöronların mevcut veya olası özelliklerinin onları açıklamak için dahil olduğu karmaşık bir deneysel veri seti hakkında düşünmeye izin veriyor .
Kaç tane hafıza deposu var ve bunlar ne için?
Beynin çeşitli bölümlerinin yok edilmesiyle ilgili yukarıda açıklanan deneylerden, iki bellek deposu olduğu sonucu çıkar - Chi X'. X deposunun, lPMGP'nin önemli bir parçası olduğu bir sistemde veya hatta lPMGP'nin kendisinde bulunduğuna dair kanıt Bölüm 1'de verilmiştir. 7.1, bölüm içinde. 5 ve 6 ve ayrıca Bölüm'de tekrar ele alınacaktır. 10. X'in lokalizasyonu, eğer bu deponun bir lokalizasyonu varsa, henüz bilinmemektedir, ancak X'in Salzen ve diğerleri (1975, 1978) tarafından keşfedilen lateral ön beyin ile ilişkili olması muhtemeldir. Bununla birlikte, bu bölgenin X' fonksiyonlarına katılımı, olasılıklardan yalnızca biridir, çünkü aynısı , WMCG'nin iletişim gönderdiği herhangi bir bölge ile ilgili olarak varsayılabilir. Bu bölgelerden biri olan yükseltilmiş paleostriatum (paleostriatum boostatum; s. 273), güvercinlerde öğrenme (Mitchell ve Hali, 1984) ve tavuklarda damgalama (Davies ve diğerleri, 1985; bkz. bölüm 9.3) için önemli olan bir kompleksin parçasıdır. . Bu çalışmalar beynin sol ve sağ hemisferleri arasında ayrım yapmadı. McCabe ve Rose ( 1985), pasif kaçınmayı geliştirirken, paleostriatal çekirdek kompleksini içeren önbeyin tabanının sol kısmından ziyade sağ kısmında metabolik kaymalar bulmuşlardır (s. 143). Bu varsayılan sistemin doğrudan deneysel analizinin yerini alamayan olası bir X' yerelleştirmesi.
Mevcut veriler, X ve X'in bağımsız depolama sistemleri olduğunu göstermektedir. lPMWG'nin sinyallerin X'e transferine "izin verdiği" varsayılabilir; bununla birlikte, LPMG'nin kendisinin X' içinde herhangi bir spesifik bilgi taşımasına gerek yoktur ve Şekil 1'de gösterilen şemada kendisine böyle bir işlev atanmamıştır. 7.9. Bilgi, geçici depolamadan X'e aktarılır . Bu şemaya göre, X ve X' , en azından yukarıda açıklanan deneyler
sırasında izleri kararlı bir şekilde depolayan paralel depolardır.

Resim: 7.10. Bir civcivin damgalamadan önce ve sonra yaklaşma ve kaçınma tepkilerini açıklayan basit bir algoritma .
Birden çok kalıcı iz deposu fikri yeni değildir (Baddeley, 1976; Warrington, 1979). Belki de daha ilginç olanı, bu sistemlerin hangi işlevleri yerine getirdiği sorusudur. Öğrenmeden önce civcivler çeşitli nesnelere yaklaşır. Bir nesneye maruz kalmanın bir sonucu olarak, tavuklar tepkilerini ona yönlendirirler. Eğitimden sonra, civcivlerin yaklaşmasına neden olan nesneler kümesi daralır: tanıdık bir nesne ve ona benzeyen uyaranlar, bir yaklaşma tepkisine neden olabilir ve benzer olmayan nesnelerden kaçınılır. Bu davranış basit yasalara uyuyor gibi görünüyor. Eğitimden önce kural şudur: “boyut, renk, kontrast, hareket hızı vb. Öğrenme, artık iki bileşeni olan bu kuralı değiştirir : 1) tanıdık bir nesneye veya ona benzeyen bir nesneye yaklaşın; 2) Ona benzemeyen nesnelerden kaçının . Bu davranış için basit bir algoritma Şekil 1'de gösterilmektedir. 7.10. Bu tür kuralların oluşturulması, "referans" belleğin bir parçası olmalıdır (Honig, 1978) ve sinirsel mekanizmaları çok basit olabilir. Bu kuralların işleyişini sağlayabilen böyle bir varsayımsal mekanizmanın bir örneği , Şekil 1'de gösterilmektedir. 7.11. Bu şema inşa edilmiştir
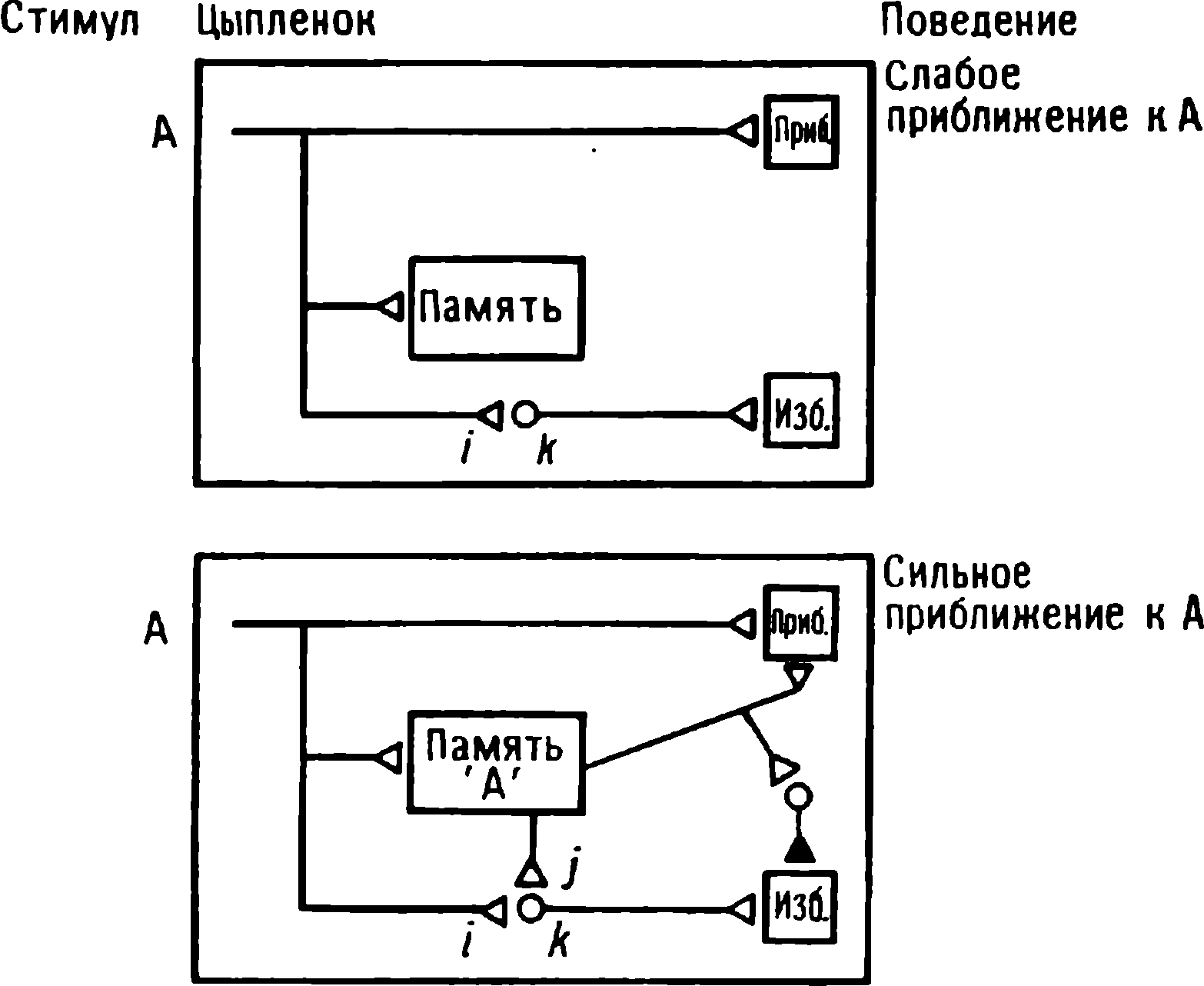
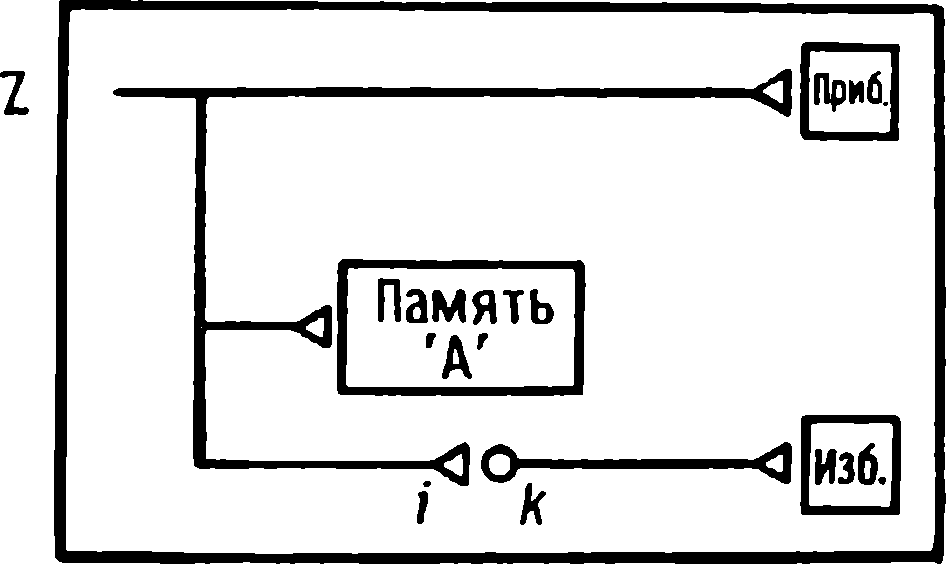
Resim: 7.11. şema
yaklaşma ve kaçınma tepkileri, tavuk sinir sistemi tarafından temsil edilir.* Büyük dikdörtgenin içindeki şekiller, belirli işlevlere sahip nöral yapıları temsil eder. yaklaşık - Yaklaşma kontrol sistemi, İzb. - kaçınma. Ûk - sinaps : eğitimsiz tavukların eksik olduğu iletim. Beyaz üçgenler uyarıcı sinapslardır, siyah üçgenler engelleyicidir. /. Eğitimsiz Tavuk, A uyarıcısına maruz kalır. İz deposunda A uyarıcısının temsili yoktur ve buna bağlı olarak, bellek sisteminden yaklaşma sistemine bir çıkış sinyali yoktur. Yaklaşma sistemine doğrudan bağlantı , kaçınma sistemine olan bağlantıdan daha verimli bir iletime sahiptir.
Uyaran A'nın kullanımıyla öğrendikten sonra. Bu uyaran sunulduğunda, hafıza sistemindeki nöral temsili heyecanlanır. Sonuç olarak, çıktısında, yaklaşım sistemini güçlü bir şekilde heyecanlandıran bir sinyal ortaya çıkar. Öğrenmede, bu sinyali yükseltmek i/k sinapsının etkinliğini artırır. Öğrendikten sonra A gösterilirken bu sinaps aracılığıyla kaçınma sistemi de devreye girer. Bununla birlikte, hafıza sisteminden gelen bir sinyalle uyarılan engelleyici bağlantı, kaçınma sistemine giden uyarıcı giriş sinyalini bloke eder. Bir civciv bir uyaran için damgalanmışsa
uyaran Z'yi gösterir, sonra bundan kaçınır. Daha ayrıntılı açıklamalar için metne bakın . Şekil l'deki ile aynı şekilde. 6.6, E, motivasyon sistemini çıkarmasına ve kaçınma tepkisini kontrol eden bir sistem eklemesine rağmen . Baskıya neden olabilecek nesneler (A - Z) , izlerin depolanmasının yanı sıra yaklaşma ve kaçınma reaksiyonlarının düzenleme sistemlerine yol açan nöral yolları uyarır. Eğitimsiz bir tavukta (Şekil 7.11, 1), bu nesnelerden herhangi biri, örneğin A, yaklaşma sistemini kaçınma sisteminden daha güçlü bir şekilde harekete geçirir . Bu nesnenin devam eden etkisinin bir sonucu olarak, hafızada sinirsel temsili oluşur . Bu nesne eğitimden sonra sunulduğunda (Şekil 7.11, //), bu temsil etkinleştirilir, depolama sisteminden ek bir sinyal gelir ve civciv nesneye aktif olarak yaklaşır (daha ayrıntılı tartışma için bkz. Bölüm 6.4). Şimdi, A nesnesi için güçlü bir tercih oluşturan civcive Z nesnesi gösterilirse , civciv bundan kaçınır (Şekil 7.11, ///). Bu davranış, Z'ye zayıf bir şekilde yaklaşan eğitimsiz bir civcivin davranışından keskin bir şekilde farklıdır.Bu değişiklik, örneğin, eğitimsiz kuşlarda i/k'yi temas halinde tuttuğu varsayılarak açıklanabilir . ama verimsiz . A'nın sunumu ile öğrenme sürecinde, hafıza sisteminden gelen sinyaller / yolu boyunca k hücresine gider (Şekil 7.11, II). k hücresine iki aktif giriş (i ve /) toplanarak boşalmasına neden olur. i/k temasının pre- ve postsinaptik elemanlarında eş zamanlı aktivitenin meydana gelmesi, bu sinapsta iletim etkinliğinde geri dönülemez bir artışa yol açabilir (Hebb, 1949). A'nın uzun bir sunumundan sonra Z göründüğünde , tıpkı eğitimsiz bir kuşta olduğu gibi, yaklaşma sistemi zayıf bir şekilde etkinleştirilir. A nesnesinin nöral temsilinin bulunduğu bellekte saklanan bilgi ile Z nesnesinin neden olduğu afferent sinyaller arasında eşleşme yoktur. Böylece hafıza sisteminden yakınlık sistemine sinyal alınmaz. Bununla birlikte, sinaps i/k boyunca iletim etkili olmaya devam eder ve kaçınma sistemi oldukça uyarılır. Böyle bir sistem, bir referans bellek kuralının veya yaklaşma ve kaçınma izinin uygulanmasını sağlayabilir, ancak böyle bir mekanizmanın gerçek sinir sisteminde çalışıp çalışmadığı bilinmemektedir . Yukarıda bahsedildiği gibi (s. 141), gelen sinyali depolanan bilgilerle karşılaştırma mekanizması da hala tartışma konusudur.
Bölümde tartışılan veriler. Şekil 7.1, hem ILMCH ile ilişkili sistem X'in hem de sistem X'in tanımayı ve referans belleği desteklediğini ileri sürmektedir. Bu bakımdan benzerler; ancak aralarında bazı farklar olması da mümkündür.
Büyük olasılıkla, bu iki sistemdeki depolama hacimleri farklıdır. X' daha büyük bir hacme sahipse, içinde depolanan bilgiler , sonraki deneyimlerin etkisi altında değişerek veya genişleyerek X'e göre yeni gereksinimlerle daha etkili bir şekilde uyumlu hale getirilebilir ve ayrıca örneğin deneyim aktarılırken daha esnek bir şekilde kullanılabilir. bir durumdan diğerine problem çözmede (Polt, 1969). Bölümde tekrar tartışılacak olan bu konular. 12.1.1 kesinlikle deneysel olarak çözülebilir.
X ve X' rolü sorusunu gündeme getirmek meşruysa, sPMVP ile ilişkili olduğu iddia edilen geçici depolamanın işlevini açıklığa kavuşturmak da meşrudur. Bu mağazadaki bilgiler belirli bir davranışsal amaç için kullanılabilir mi? Henüz bir cevabı olmasa da bu soru sorulmayı hak ediyor. Bir olasılık, orada depolanan bilgilerin belirli bir görevi veya görev dizisini gerçekleştirmek için kullanılmasıdır. Bu tür depolama "işleyen bellek" olarak bilinir (Baddeley, 1976; Honig, 1978; Olton ve diğerleri, 1979; Olton ve Feustle, 1981; ayrıca bkz. Konorski, 1967). Çalışma belleği , belirli bir denemenin başarıyla tamamlanması için gerekli bilgilerin deposu olarak görülür. Ancak, bir numuneden diğerine saklanmaz, çünkü bu sadece karışıklığa neden olabilir. Belirli bir görevle ilgili ayrıntılı bilgiler, çözüldüğünde silinebilse de , görevi çözerken elde edilen bazı bilgilerin saklanması mantıklı olabilir. Örneğin, özellikle tatsız veya özellikle hoş olaylarla ilgili bilgiler, çalışan bellekten uzun süreli bir depolama sistemine aktarılabilir. O zaman sPMSH çalışan bir bellek sistemi veya bunun bir parçası olabilir ve X' uzun süreli bir bellek sistemi olabilir.
Çalışan bellek kavramı, hem kuşların hem de memelilerin davranışlarının incelenmesine uygulanmıştır (Honig, 1978). Bu kavramı yiyecek arama davranışının analizinde kullanmak özellikle uygundur. Bir kuş yiyecek aradığında veya daha önce saklandığı yerde bulduğunda, kuşun az önce nerede olduğunu hatırlaması önemlidir. Bu davranış birçok kuş türünde gözlemlenmiştir (örneğin bkz. Kamil, 1978; Sherry ve diğerleri, 1981; van der Wall, 1982; Sherry, 1984). Bu verimli olabilir, ancak işleyen bellek kavramı, damgalama analizinde kullanılmamıştır. Örneğin, bir civciv annesinden ayrılırsa, ona geri dönebilmesi için nerede olduğunu hatırlaması önemlidir. Bu bilgilerin uzun süreli depolamaya aktarılması uygun değildir.
X, X' ve geçici depolamanın işlevleri hakkında burada yazılanların çoğu spekülatiftir, çünkü özellikle bu akıl yürütmenin dayandığı deneysel sonuçlar nispeten yenidir. Bu alan deneysel araştırmalar için çok verimli olmayı vaat ediyor .
Beyin hasarı, motivasyon ve uyaran-tepki bağlantısını sağlayan yollar
Yukarıdakiler, PMVH'deki biyokimyasal değişiklikleri ve hasarının etkilerini uyanıklık ve dikkat düzeyindeki değişikliklere veya duyusal ve motor işlevlerdeki değişikliklere değil de hafıza işlevinin belirli yönlerine bağlamamızın nedenlerinden bazılarıdır (bkz. 3 ve 5 ve ayrıca bölüm 6.1, 6.2 ve 8.1). Bu bölümde anlatılan deneyler aynı konuyla, ama özellikle motivasyon sorunuyla ilgilidir. PMVP'deki hasar, civcivi tanıdık bir nesneye yaklaşmak için gereken motivasyonu kaybedecek şekilde etkiler mi? Sıralı yaralanma denemelerinde (Bölüm 7.1), test 2'de tercihleri ölçüldüğünde her iki tavuk grubunda da iki taraflı MAH lezyonları vardı . Ancak, bir grup rastgele seçimle, diğeri ise tam nesne tanımayla karakterize edildi. Bu sonucu motivasyon sistemine verilen zararla açıklamak zordur. Bunu motivasyonel bir bozukluk olarak açıklamanın bir diğer zorluğu da , civcivlerin eğitimden sonraki gün IPMCH ve pPMCH çıkarıldığında tanıdık nesneyi tercih etmeye devam etmeleridir (Şekil 7.6b); eğitimden 3 saat sonra bilateral cerrahi yapıldığında durum böyle değildi (bkz. Şekil 5.6). Diğer sonuçlar da bu açıklamayı desteklememektedir. Bir sonraki bölümde gösterileceği gibi, PMSH'ye verilen hasar tavukların davranışına yansırsa, bu etki yalnızca kutunun sunumu ile eğitilenlerde güçlü bir şekilde , ancak eğitim verilenlerde zayıf olarak telaffuz edilir. doldurulmuş tavuk sunumu. PMVG hasarı motivasyonu etkiliyorsa, bu yalnızca bir grup tavukta meydana gelir. Bu sonuçlar , yukarıda belirtilen bölümlerde açıklananlarla birlikte , PMA hasarının tercih üzerindeki etkisinin motivasyon durumundaki değişikliklerle açıklanamayacağını göstermektedir.
Bu bölümde açıklanan sonuçlardan bazıları başka bir genel soruna da değinmektedir. Baskıya uygun bir nesneye tepki veren duyusal nöronların , yaklaşma tepkisini düzenleyen nöronlarla bağlantısını tek bir düz çizgi olarak tasavvur etmek mümkün müdür ? Durum buysa, nesnenin sunumu uyaran-tepki bağlantısını güçlendirmeli ve basit bir refleks arkının oluşumunu desteklemelidir. Öğrenmeden önce bu arkı kırmak, böyle bir pekiştirmenin ve dolayısıyla bir becerinin oluşmasını imkansız hale getirecektir . Bağlantı geliştirme tamamlandıktan sonra arkı kırmak, duyusal ve motor nöronların bağlantısını kesecek ve tanıdık nesne, yaklaşma tepkisini ortaya çıkaramayacaktı. Bu davranış, bir hafıza kusuru olarak yorumlanabilir. SMG böyle bir arkın bir bileşeni olsaydı, hatta depolama işlevine sahip özel bir bileşen olsaydı, hasarının etkisi mutlaka herhangi bir ark bağlantısının hasarından farklı olmazdı. Ancak bu fikir doğruysa, o zaman PMSH'deki hasar her zaman tanıdık nesne için edinilen tercihi bozmalıdır , ki durum böyle değildir (bkz. Şekil 7.3 ve 7.6b ). Bu nedenle, PMSH basitçe bir uyarıcı ve bir tepkiyi birbirine bağlayan bir arkın parçası olarak düşünülemez. Böyle bir ark varsa, SMVP paralel, bağımsız bir yol oluşturmalıdır (bkz. Şekil 6.6 ve 7.11). PMSH'ye verilen hasarın çağrışımsal öğrenme üzerindeki etkilerine ilişkin bir çalışmanın sonuçları (McCabe ve diğerleri, 1982) aynı şeyi göstermektedir: iki görsel nesneden birine seçici bir yaklaşım yanıtının üretilmesi, görsel nesnenin hasar görmesi ile bozulmaz. Doğru tepki pekiştirildiğinde PMSH. PMVG, afferent ve efferent nöronlar arasındaki basit bir doğrusal yolun parçası olsaydı, böyle bir edimsel yanıtın üretilmesi imkansız olurdu .
Özet
, damgalama işlemi sırasında sol ve sağ PMVP'nin fonksiyonlarının interhemisferik asimetrisine işaret eden verileri sunar . LPMVG'nin bir depolama işlevine sahip olduğu ve kendisinin bir iz deposu olabileceği varsayılmaktadır. Ancak, LPWG tek depo değildir. Özel koşullar altında , önceden geliştirilen tercihi bozmadan her iki PMVP'yi de kaldırmak mümkündür . Bu sonuçlara dayanarak, X' etiketli başka bir depo olduğu sonucuna varılmıştır. Bu kasa , geçici bir kasa görevi görebilen rMPVG'nin kontrolü altında yavaş yavaş oluşuyor gibi görünüyor . İki PMHG'nin etkileşimi ve varsayılan depolama X' için bir nöral model önerilmiştir. İnsanlarda olduğu gibi diğer hayvanlarda da çoklu hafıza sistemlerinin varlığına ilişkin veriler sunulmaktadır . Bu hafıza sistemlerinin tavuklardaki olası işlevleri tartışılmıştır.
EDEBİYAT
Baddeley A. D. Hafıza psikolojisi. Haagre ve Row, New York, 1976. oradley P, Davies DC, Horn G. Evcil civcivin (Gallus domesticus) hyperstriatum ventrale bağlantıları, J. Anat., 1985, 140, 577-589.
Chen CLC, Dionne FT, Roberts JL Dopaminerjik bileşikler tarafından sıçan hipofizindeki mRNA seviyelerinde proopiyomelanokortinin düzenlenmesi, Proc. nat. Acad. bilim ABD, 1983, 80, 2211-2215.
Cipolla-Neto J., Horn G., McCabe BJ Hemispherio asimetri ve damgalama: sıralı lezyonların hyperstriatum ventrale'ye etkisi, Exp. Brain Res., 1982, 48, 22-27.
Daley JE, McCabe BJ, Horn G. Evcil civciv Behav'da baskıdan sonra bilgi depolama mekanizmaları. Brain Res., 1987 (baskıda).
Davies DC, Horn G „ McCabe BJ Noradrenalin ve öğrenme: noradrenerjik nörotoksin DSP4'ün evcil civciv Behav'da damgalama üzerindeki etkileri. Neurosci., 1985, 99, 652-660.
Dennis M., Kohn B. Serebral hemidekortikasyon sonrası infantil hemiplejiklerde sözdiziminin anlaşılması: sol hemisfer üstünlüğü, Brain and Language, 1975, 2, 472-482.
Hebb D. 0. Davranışın organizasyonu, John Wiley, New York, 1949.
Nösaen, H. Çocuklarda edinilmiş afazi ve hemisferik fonksiyonel uzmanlaşmanın ontogenezi, Brain and Language, 1976, 3, 114-134.
Honig WK Güvercinlerde çalışma belleği çalışmaları. İçinde: Hayvan davranışında bilişsel süreçler (editörler SH Hulse, H. Fowler ve WK Honig), 1978, s. 211-248, Eribaum, Hillsdale, NJ
Horn G., McCabe BJ, Cipolla-Neto J. Yerli civcivde baskı: hiperstriatum ventrale'nin her iki tarafının edinme ve tutmadaki rolü, Exp. Brain Res., 1983, 53, 91-98.
Kamil AC Nektarla beslenen bir kuş olan Amakikhi (Loxops virens), J. comp. tarafından sistematik yiyecek arama. fizik. Psychol., 1978, 92, 388-396.
Konorski J. Beynin bütünleştirici aktivitesi. Chicago Press Üniversitesi, Chicago , 1967.
Levy J. İnsan beyninin yanal uzmanlaşmasının evrimi için olası temel, Nature, Londra., 1969, 224, 614-615.
Levy J. Bilateral asimetrinin psikobiyolojik sonuçları. İçinde: İnsan beyninde yarım küre işlevi (eds. SJ Dimond ve JG Beaumont), 1974, s. 121-183, Elek (Bilimsel Kitaplar), Londra.
Marr D. Serebral korteks için bir teori, Proc. R. Soc., 1970, B176, 161-234.
Marr D. Basit bellek: Archicortex için bir teori, Phil. Trans. R. Soc., 1971, 262, 23-81.
McCabe BJ, Cipolla-Neto J., Horn G., Bateson PPG Baskıdan sonra civcivin hiperstriatum ventralinde yer alan bilateral lezyonların amnezik etkileri , Exp. Brain Res., 1982, 48, 13-21.
McCabe BJ, Hunt S. P. Yerli civcivin telensefalonundaki peptitler ve biyojenik aminler. Bir immünositokimyasal çalışma, Neurosci. Lett., 1983, Ek. 14, S242.
McCabe N Rose SPR Pasif kaçınma eğitimi in vitro fukoz birleşmesini artırır, Neurochem Res., 1985, 10, 1083-1095.
Mitchell JA, Hali G. Güvercinlerde paleostriatal lezyonlar ve enstrümantal öğrenme, Quart. J. exp. Psychol., 1984, 36B, 93-117,
Moscovitch M. Dil işlevlerinin lateralizasyonunun gelişimi ve bunun bilişsel ve dilsel gelişimle ilişkisi: bir inceleme ve bazı teorik spekülasyonlar. İçinde: Dil gelişimi ve nörolojik teori (eds. SJ Segalowitz ve F.A. Gruber), 1977, s. 193-211, Academic Press, New York.
Hottebohm F. Kuşlarda ses öğrenimi için beyin yolları: ilk 10 yılın gözden geçirilmesi, Prog. Psychobiol. fizik. Psychol., 1980, 9, 85-124.
Nottebohm F., Stokes T. M „ Leonard CM Kanaryadaki şarkının merkezi kontrolü, Serinus canarius, J. comp. Neurol., 1976, 165, 457-486.
Olton DS, Feustle WA Mekansal olmayan çalışma için gerekli olan hipokampal fonksiyon
bellek, Exp, Bra in Res., 1981, 41, 380-389.
Olton DS, Becker JT, Handelmann GE, Hipokampus, uzay ve hafıza, Behav. Brain Sci.1979, 2, 313-365.
Polt JM Damgalama deneyiminin civcivlerde ayrımcılığı öğrenme üzerindeki etkileri, J. comp. fizik. Psychol., 1969, 69, 514-518.
Potter E., Nicolaisen AK, Ong ES, Evans RM, Rosenfeld MG Tirotropin salan hormon, RNA transkriptinde birincil prolaktin üretimini artırmak için hızlı nükleer etkiler uygular, Proc. nat. Acad. bilim ABD, 1981, 78, 6662-6666.
Salzen EA, Parker DM, Williamson AJ Evcil civcivlerde baskıyı önleyen bir ön beyin lezyonu, Exp. Brain Res., 1975, 24, 145-157.
Salzen EA, Parker DM, Williamson AJ Evcil civcivlerde ön beyin lezyonları ve imprinting retansiyonu, Exp. Brain Res., 1978, 31, 107-116.
Salzen EA, Williamson AJ, Parker DM Evcil civcivlerde ön beyin lezyonlarının doğuştan gelen ve damgalanmış renk, parlaklık ve şekil tercihleri üzerindeki etkileri, Behav. İşlem., 1979, 4, 295-313.
Sherry D. Siyah şapkalı bülbüllerle yiyecek saklama: önbelleklerin konumu ve içeriği için bellek, Anim. Behav., 1984, 32, 451-464.
Sherry DF, Krebs JR, Cowie RJ Bataklık göğüslerinde depolanan yiyeceklerin yeri için bellek, Anim. Behav., 1981, 29, 1260-1266.
Thoenen H., Kreutzberg GW (ed.) Sinir sisteminde hızlı taşımanın rolü, Neurosci. Res. programı. Bull., 1981, 20, 1-138.
Trevarthen C. Dil için serebral mekanizmaların gelişimi. İçinde: Dil, okuma ve hecelemenin nöropsikolojisi (ed. U. Kirk), 1983, s. 47-82. Academic Press, Londra.
Van der Wall SB Clark's Nutcracker, Anim'de önbellek kurtarmanın deneysel bir analizi. Behav., 1982, 30, 84-94.
Warrington EK Çoklu bellek sistemleri için nöropsikolojik kanıtlar. İçinde: Beyin ve akıl, Ciba Vakfı Sempozyumu no. 69 (Yeni Seri), 1979, s. 153-166. Alıntı Medica, Amsterdam.
Zigmond RE, Chalazonitis A. Preganglionik sinir stimülasyonunun sıçan üst servikal ganglionundaki tirozin hidroksilaz aktivitesi üzerindeki uzun vadeli etkileri, Brain Res., 1979, 164, 137-152.
BÖLÜM 8
EĞİLİM VE TERCİH
Tavuklar ve ördek yavruları çok çeşitli görsel uyaranları takip edebilir, ancak bu çeşitlilik sınırsız değildir ve bazı nesneleri takip etmede diğerlerinden çok daha etkilidir. Fabricius ve Boyd ( 1952/53) , bir kibrit kutusundan daha büyük olmayan nesnelerin, ördek yavruları tarafından yiyecek olarak görmezden gelindiğini veya takip edildiğini ve damgalama uyaranları olarak bu nesnelerin nispeten etkisiz olduğunu buldular. Schaefer ve Hess ( 1959), tavuklarda aşağıdaki tepkinin mavi ve kırmızı uyaranlar tarafından sarı veya beyaz uyaranlara göre daha kolay ortaya çıkarıldığını gösterirken, Kovach ( 1971) yeşilin yaklaşma tepkisini oluşturmada diğerlerinden daha az etkili olduğunu buldu. Boyut , şekil, kontrast, hareketin varlığı veya yokluğu ve hareketin yönü, bir yaklaşma tepkisini ortaya çıkarmadaki etkinliğini belirleyen bir uyaranın parametreleridir ve bu liste ayrıntılı olmaktan uzaktır (bkz . Sluckin , 1972; Kovach, 1983).
Çoğu deneysel çalışmada, civcivlerin yaklaşmasına neden olan uyaranların özellikleri incelenirken, doğal ebeveynlerden daha çok yapay nesneler kullanılmaktadır. Bunun faydaları yeterince açıktır. Yapay nesneler kullanıldığında, parametreleri çeşitli şekillerde değiştirilebilir, bu canlı kuşlarda imkansızdır ; ayrıca yapay bir nesne, ebeveyninin aksine davranışını değiştiremez. Aynı zamanda, yöntemin bir dezavantajı vardır , çünkü bazı kuş türlerinin civcivlerinin kendi türlerinin bireylerinin görsel işaretlerini öğrenme sürecinde herhangi bir belirli özelliğin olup olmadığı sorusunu netleştirmeye izin vermez. . Pek çok deneyde , yaklaşma tepkisini geliştirmek için doldurulmuş hayvanlar veya onların basit modelleri kullanıldı, bu nedenle sorulan soruyu netleştirmek için bu çalışmalara dönmek gerekiyor. Aynı zamanda, hiçbir hayvan modelinin yaşayan bir hayvanın aynısı olmadığı gerçeğini de unutmamak gerekir . Buna göre, tür tanımlama sorunu açıklığa kavuşturulurken, bu çalışmaların sonuçları dikkatle yorumlanmalıdır.
Johnston ve Gottlieb ( 1981) bu sorunu evcil (Pekin) ördek yavrularında ve yabani yeşilbaş ördek yavrularında (Anas platyrhynchos) araştırdılar. Ördek yavruları 24 saatlik olduklarında, 20 dakika boyunca bir dizi nesneyi takip etmelerine izin verildi : doldurulmuş bir yabani tavuk, bir veya iki su kuşu türünün doldurulmuş hayvanları ve bir veya iki geometrik şekil . Daha sonra ördek yavrularına , eğitimde kullanılan modeli ve diğer modellerden birini aynı anda sundukları bir seçim testi verildi . Bu deneylerin ana sonucu , ördek yavrularının kendi türlerinin üyelerini takip etmeye yatkınlıkları varmış gibi davranmalarıydı. Bu yatkınlık, ördek yavrularına geometrik şekilleri takip etmeyi öğreterek değiştirilebilir; ördek yavrularına su kuşu türlerinden biri gösterilirse kısmen üstesinden gelinebilir, ancak tamamen ortadan kaldırılamaz. Bu sonuçları yorumlarken çok dikkatli davranan ve bunları 'yatkınlık' ile açıklamayı teklif etmeyen Johnston ve Gottlieb, çalışmalarını ' tür tanımlama sorununa basit bir yaklaşım'a karşı bir uyarı ile sonlandırdı . Özellikle beyin hasarının damgalama üzerindeki etkisini incelerken, değiştirilmiş bir biçimde de olsa, bu sorunun ortaya çıkmasıyla bağlantılı olarak onlara katılmamak zordur .
Beyin hasarı ve tercih
Beyin hasarının damgalama üzerindeki etkilerini incelemek için tasarlanan deneylerin her birinde, eğitimde kullanılan nesneler ya dönen, yanıp sönen kırmızı bir kutu ya da doldurulmuş bir banka tavuğuydu ( sırasıyla Şekil 2.2, A ve B). Doldurulmuş bir yabani tavuk, kendi ekseni etrafında ya sallandı ya da döndü. Hasar, eğitimden önce veya sonra, ayrıca eğitimden önce ve sonra uygulandı (Bölüm 5 ve ayrıca Bölüm 7.1). Nesne sunulduktan sonra, tavuğun bu nesneyi tercih etme gücü , diğer nesneleri tercih etmesiyle karşılaştırılarak ölçüldü . Yeni nesne, civcivin eğitim sırasında görmediği şeye bağlı olarak bir kutu veya doldurulmuş bir hayvandı . Test sırasında civcivlere sırasıyla kutu ve doldurulmuş hayvan farklı bir sırayla sunuldu. Her çalışmada, PMVH'si hasar görmüş en az iki tavuk grubu ve sahte olarak çalıştırılan bir kontrol grubu kullanıldı. Deneylerin hiçbirinde kutu eğitimli ve kukla eğitimli tercih puanları arasında önemli bir fark bulunmadı , ancak bu özellikle istatistiksel yöntemler kullanılarak tespit edilmeye çalışıldı. Ancak, tüm deneyler için elde edilen ortalama tercih puanları

Resim: 8.1. PMVP'ye verilen hasardan sonra tercih puanlarının düştüğü deneylerden elde edilen verilerin özeti . PMVP yaralanması olan tavuklar ve sahte ameliyat edilen tavuklar için göstergeler, eğitim sırasında sunulan uyarana bağlı olarak gruplandırılır. Gösterilenler, bir kutu (A) ve doldurulmuş tavuk (B) kullanılarak eğitilmiş tavuklar için ortalama değerler ve standart hatalardır . Standart hatalar, rastgele 50 düzeyine sahip göstergeleri karşılaştırmak için de kullanılabilir. Tablodan veriler. 8.1.
birlikte ele alındığında (Tablo 8.1), kutuyla eğitilen beyin hasarlı civcivlerin puanlarının kuklayla eğitilen civcivlere göre sürekli olarak daha düşük olduğu bulundu (Hoth ve McCabe, 1984).
Tablo 1'de yer alan ortalama göstergelerin hesaplandığı ilk tercih göstergelerinin istatistiksel bir analizi yapılmıştır. 8.1. Kutu eğitimli sahte çalıştırılan kontrol civcivlerinin (78.68) genel ortalama tercih puanı (Şekil 8.1), kukla eğitimli kontrol civcivininkine (83.49) çok yakındı . Kutuyla eğitilen beyin hasarlı civcivler , tanıdık uyarıcıyı tercih etmediler: bu civcivler için genel ortalama tercih puanı 50.68 idi. Buna karşılık, doldurulmuş bir tavukla eğitilen deneysel tavuklar , tanıdık uyaranı tercih ettiler. Genel ortalama tercih puanları (72.19), kutu eğitimli deney grubu için hem rastgele 50 seviyesinden hem de genel ortalamadan önemli ölçüde farklıydı. Bu sonuçlar, PMVP'deki hasarın eğitimli tavuklarda tercihi etkilemediği anlamına mı geliyor?
Tablo 8.1. Hasarlı PMA ve Sahte Kontrollere Sahip Civcivlerde Ortalama Tercih Puanları (Horn ve McCabe, 1984)
Veriler, eğitim sırasında farklı uyaranlarla sunulan tavuk grupları için verilmiştir. Parantez içindeki rakamlar gruptaki civciv sayısını göstermektedir. Sonuçlar, tercih puanlarının PMVG'ye verilen hasarla kötüleştiği tüm deneylerden alınmıştır. Tabloda gösterilen standart hatalar, genel ortalamaları rastgele 50 düzeyinde karşılaştırmak için kullanılır. Ortalamalar arasındaki karşılaştırma için, orijinal verilerin analizinden elde edilen standart hatalar kullanılır (Horn ve ark . .67 ). ; P<0.001). Grubun karşılıklı etkisi vardır.
hasar ve kontrol) ve öğrenme koşulları (G 1>181 =5.92; P<0.025). Doldurulmuş tavuğun değiştirilmesiyle eğitilen kontrol civcivlerinin genel ortalama puanı önemli ölçüde farklıdır: i) 50'den (PC0.001), 2) kutuyla eğitildiğim hasarlı civcivlerin genel ortalama puanından (P<0.0()) 1) ve 3) doldurulmuş tavuğun değiştirilmesi ile eğitilmiş hasarlı PMCH'li grup için genel ortalamanın (P<0.01). Bu durumda, farklı deney dizilerindeki tercih göstergeleri,
♦ Önce sağ PMVG'nin, sonra solun hasar gördüğü civcivler (bölüm 7.1).
Resim: 8.2. Wulst hasarının tercih tepkisi üzerindeki etkisi. Hasarlı kanatlılar (W) ve kontrol grubu (LO) için veriler, Tabloda gösterilen son üç deneyden alınmıştır. 8.1. Wulst lezyonları , kutu (A) ve doldurulmuş tavuk (B) ile eğitilmiş civcivlerin ortalama tercih puanlarını önemli ölçüde etkilemedi .
korkuluk kullanmak? Bu uyaranla eğitilen kontrol civcivlerinin genel ortalama tercih puanı 83.49'du. Bu gösterge, aynı koşullar altında eğitilmiş beyin hasarı olan tavuklardan önemli ölçüde daha yüksektir (P<0.01). Bu nedenle, doldurulmuş bir tavukla eğitilen deneysel tavuklar , yanıtta küçük ama istatistiksel olarak anlamlı bir düşüş gösterdi.
PMVG'si hasar görmüş kanatlılarda tercih puanlarındaki düşüş, kısmen beyin hasarının bazı spesifik olmayan etkilerinden kaynaklanıyor olabilir. Ancak, durum pek öyle değil. Tabloda gösterilen son üç deney. 8.1 , Wulst hasarına sahip ek kontrol gruplarını içeriyordu. Kutu ve doldurulmuş tavukla eğitilen bu civcivlerin ortalama tercih puanları kontrol grubundan önemli ölçüde farklı değildi (Şekil 8.2).
Tercih indeksindeki azalma farklı şekillerde oluşabileceğinden (s. 104), Tablo 1'de listelenen tüm deneylerde iki nesneye yaklaşma tepkilerini inceledik. 8.1. Tek önemli etki, onlara bir kutu verilerek eğitilen PMVG'si zarar görmüş civcivlerde bulundu. Nesneye, sahte olarak çalıştırılanlardan önemli ölçüde daha az yaklaştılar (P < 0.05), yani hasar etkisi, alışılmadık nesnelerden duyulan korkuyu azaltmadı . Durum böyle olsaydı, beyin hasarı olan tavukların yeni nesneye kontrol tavuklarından daha aktif yaklaşması gerekirdi. Gerçekte, kendilerine sunulan nesneyi bilmiyormuş gibi, yani hafızaları bozulmuş gibi davranıyorlardı.
Sonuçlar tabloda sunulmuştur. 8.1, dört deneyin hepsinde şaşırtıcı bir şekilde sabittir . Onlara bir kutu veya doldurulmuş tavuk sunularak eğitilen sahte ameliyat civcivleri , yüksek tercih puanları elde etti ve iki genel ortalama puan neredeyse aynıydı. PMVG hasarının tanıdık bir uyaran tercihini bozduğu deneylerde, eğitimde kullanılan nesnenin doğasına bakılmaksızın performans azaldı, ancak bu etki kutu eğitimi ile telaffuz edildi ve doldurulmuş bir hayvanla nispeten zayıftı.
Belki de bu sonuçların en basit açıklaması, iki nesne arasındaki karmaşıklık farkında yatmaktadır, çünkü doldurulmuş tavuk, kırmızı kutuya kıyasla renk ve uzamsal kontrast gibi birçok özellik hakkında önemli ölçüde daha zengindir. Bu farklılıklar nedeniyle, dönen bir kutu hakkındaki bilgiler depoda nispeten az "yer" kaplayabilir ve SMCG'nin içinde bulunan sinir ağlarında yer alabilir. Buna karşılık, doldurulmuş bir tavuk hakkındaki bilgi nispeten büyük miktarda depolama alanı kaplayabilir ve karşılık gelen nöron sistemleri beyinde daha geniş bir şekilde dağılabilir. Bu durumda PMVG'nin zarar görmesi kutunun tanıma sistemini bozacak ancak doldurulmuş tavuğun tanınmasını bozmaya yetmeyecektir.
Sonuçların tamamen farklı bir açıklaması, doldurulmuş tavuğun olası biyolojik öneminden geliyor. SMVP'deki hasarın, bu nesnenin tercih edilmesi üzerinde (küçük de olsa) bir etkisi vardır. Böyle bir sonuç, doldurulmuş tavuğu tanıma sisteminin önemli bir kısmının PMVG kapsamı dışında kaldığı anlamına gelebilir. Bu nesneyi tanımak için biri PMVP'de ve diğeri dışında olmak üzere iki sistem olması ve bu sistemlerin farklı işlevleri olması mümkün mü? Durum böyleyse, aynı türün bireylerinin bazı ortak özelliklerini tanımak için SMGS dışında bir sisteme ihtiyaç duyulmuyor mu ve aynı türün temsilcilerinin bireysel özelliklerini tanımak için SMGS'de bir sisteme ihtiyaç duyuluyor, örneğin, anneler, rengi, işaretleri, tüyleri vb. hakkında bilgi depolamak için? PMVP'ye verilen hasardan sonra kırmızı kutu tercihinin güçlü bir şekilde ihlali, bu alanın yalnızca türün bireysel temsilcilerinin değil, aynı zamanda diğer nesnelerin de tanınmasında yer aldığını göstermektedir. Bu düşünceler , spekülatif doğaları akılda tutularak özetlemenin mantıklı olduğu ilginç varsayımlara yol açar :
hem aynı türün bireylerinin hem de diğer nesnelerin görsel özellikleri. Öğrenmeden önce, nesnenin nöral temsilinin - "iz" veya "engram" - olmadığı ve bunların oluşumunun nesnenin gösterilmesiyle eğitim gerektirdiği varsayılır; bu son söz aşırı derecede bilgiççe görünebilir, ancak Bölüm'den görüleceği gibi değildir. 8.2; 2) SMVG'nin dışında, aynı cins veya türden bireylerin belirli ortak özelliklerini tanımaya yönelik bir sistem vardır.
Davranışsal Tercih Analizi
yukarıda bahsedilen "çifte tanıma sistemi"nden ziyade "uyarıcı karmaşıklığı" (s. 179) terimleriyle açıklamak daha kolaydır . Ek olarak, ikili tanıma sistemi kavramı doğruysa , uzun yıllar bozulmamış tavukları inceledikten sonra, davranışlarında doldurulmuş tavuk için daha güçlü bir tercih veya ona yaklaşma konusunda seçici bir eğilim göstermemiş olmamız gariptir. Böyle bir eğilim meydana gelebilir, ancak beyin hasarı deneylerinin biri hariç hepsinde kullanılan sıralı test yeterince hassas değilse fark edilmeyebilir. Gerçekten de, bu testin, nesnelerin aynı anda gösterilmesiyle yapılan seçim testinden çok daha az duyarlı olduğuna inanmak için sebepler var (Şekil 3.2, B, C). Ardışık testte, yaklaşma tepkisi tekerleğin devir sayısıyla ölçülür. Çarkın çevresi 94 cm olduğu için civcivin çarkın hareketinin kaydedilebilmesi için mesafenin en az yarısı yani 47 cm koşması gerekir . Buna karşılık, eşzamanlı seçim testinde platform hareketi en yakın 1 cm'ye kadar ölçülür, 1 cm'lik bir platform kayması, civcivin çarkın çevresinde 5,7 cm hareket etmesine karşılık gelir. Ortalama olarak civcivin 1 cm'lik bir platform hareketinin kaydedilmesi için bu mesafenin yarısını koşması yeterlidir. Bu nedenle, sıralı testteki ölçü birimi , civcivin çarkın çevresinde 47 cm'den fazla, ancak 141 cm'den az hareketine karşılık gelir; eş zamanlı seçim testinde karşılık gelen hareket 2,85 cm'den fazla, ancak 8,5 cm'den küçüktür, bu nedenle, diğer şeyler eşit olduğunda, eş zamanlı test, sıralı testten 16 kat daha hassastır. Ek olarak, uyaranları eşzamanlı sunumlarıyla ayırt etme görevinin sıralı sunumdan daha kolay olduğuna inanmak için sebepler vardır ( Mackintosh, 1974). Bu nedenle, eşzamanlı seçim testi daha hassastır ve tercihteki daha ince değişiklikleri tespit edebilir. Aşağıda açıklanan deneylerde, bu test, rayların uçlarında kırmızı bir kutu ve doldurulmuş bir tavukla kullanıldı.
Test 1 Test 2 O 19 23 27 2 9 51
A ■ I R—' | -С_ h
BIR - 1 * - 1 * ben
Resim: 8.3. Edinilmiş tercih ve yatkınlık çalışmasında deneysel tavukların tedavi şeması (Johnson ve diğerleri, 1985). "P" süresi boyunca civcivler 30 dakika süreyle ışığa maruz bırakıldı. 4 saat sonra, bu seriler arasında 2 saat arayla iki seri edimsel davranış eğitimi denemesi yapıldı (Şekil 6.3). Aktif pedala basarak tavukların yarısı (A) ışıklı dönen kırmızı kutuyu, geri kalanı (B) doldurulmuş tavuğu gördü. Tavukların yarısında tercih 2 saat sonra (test 1) ve geri kalanında eğitim bitiminden 24 saat sonra (test 2) ölçüldü . Tercih, eş zamanlı seçimli testlerde ölçülmüştür (Şekil 3.2, B, C).
£ 2.1. Edinilmiş tercih ve yatkınlık arasındaki etkileşim
İlk deney dizisinde (Johnson ve diğerleri, 1985), tavuklar bir edimsel öğrenme aparatında pedala basarak dönen bir doldurulmuş tavuğu veya kırmızı bir kutuyu görebildiler (Bölüm 6.2 ve Şekil 6.3). Civcivler daha sonra karanlık inkübatöre geri gönderildi. Tercih yanıtları 2 saat sonra (test 1) veya 24 saat sonra (test 2) test edildi (Şekil 8.3). Dört tavuk grubunun tümü için tercih indeksleri, Şekiller 1 ve 2'de gösterilmektedir. 8.4, A, B. 2 saat sonra test edildiğinde civcivler , ister doldurulmuş tavuk ister kırmızı kutu olsun tanıdık nesneyi tercih ettiler. Puanlar önemli ölçüde farklılık göstermedi. 24 saat sonra test edildiğinde durum değişti: Doldurulmuş tavukla eğitilen civcivlerin ortalama tercihi önemli ölçüde artarken , kırmızı kutuyu ilk görenlerinki de aynı derecede önemli ölçüde azalarak rastgele bir 50 düzeyine indi.
Testte hem doldurulmuş hayvan hem de kutu aynı anda sunulduğundan, tercihin gücü, Tavuğun önceki deneyimlerinden bağımsız olarak doldurulmuş tavuğa bir yaklaşım olarak ifade edilebilir. Doldurulmuş tavuk tercihini ifade eden ortalama göstergeler , Şek. 8.4, C, D. Bir kutu ile sunulan ve 1. testte bu nesneyi tercih eden tavuklar, doğal olarak, doldurulmuş tavuğu tercih etmekte zayıftı. Bununla birlikte, eğitimden 24 saat sonra test edilen tüm civcivler, eğitim sırasında kullanılan uyaran ne olursa olsun, 2 saat sonra test edilenlere göre doldurulmuş hayvanları önemli ölçüde daha güçlü bir şekilde tercih etti .
Daha önce nesne ile deneyim sahibi olan tavuklarda doldurulmuş tavuğun tercih edilmesindeki artış şaşırtıcı değil. Ancak, aynı ten rengine dair işaretlerin ortaya çıkması beklenmedik bir durumdu.

Resim: 8.4. Kutu (A, C) ve doldurulmuş tavuk (B, D) ile eğitilmiş civcivler için test 1 ve 2'deki ortalama tercih puanları . A ve B için tercih göstergeleri aşağıdaki formülle hesaplanmıştır:
tanıdık bir nesneye yaklaşmaya çalıştığında platformun kat ettiği mesafe )
Platform tarafından kat edilen toplam mesafe
C ve D için , A ve B'de sunulan tercih puanları doldurulmuş tavuk tercih puanlarına göre ifade edilir (bkz. Şekil 8.6 başlığı). (Johnson ve diğerleri, 1985.)
Tercih testinden önce kutuyu gören ancak doldurulmuş civcivleri görmeyen civcivlerde dentia. Takma eğitimli civcivlerdeki tercih artışı " birleştirme" ile açıklanabiliyorsa, diğer grup için de öyle kabul edilmelidir. Buna göre, bu civcivlerde tanıdık uyaran tercihi de test 2'de test 1'e göre daha yüksek olmalıdır, ancak bu gerçekleşmez. Bu kuşlarda performansın rastgele bir seviyeye düşmesi ve dolma tavuğun tercih edilmesindeki bariz artışın sadece bir yanlış yorumlama olması mümkün mü? Bu konuya açıklık getirecek verilerin elde edilmesi gerekmektedir.
Bu tür veriler, şeması Şekil l'de gösterilen deneyde elde edildi. 8.5. Tavuklar genel aydınlatmaya maruz bırakılmadan önce karanlıkta tutuldu. Daha sonra iki gruba ayrıldılar. Bir (“hafif”) grubun civcivleri (Şekil 8.5, H) , mat siyah boya ile boyanmış odalara yerleştirilmiş dönen tekerleklere birer birer yerleştirildi . Civcivler içlerinde kaldığı sürece odalar loş bir şekilde aydınlatılmıştı. Bu hücrelerde herhangi bir nesne yoktu. Bu deneyden sonra, civcivler karanlık inkübatörün ayrı bölmelerine geri konuldu . Diğer civciv grubu, karanlık bir kuluçka makinesinde ("karanlık" grup) her zaman izole kaldı (Şekil 8.5, B). Vasıtasıyla
Test 1 Test 2
19 23 29 51 _
l ben r ben — < 1
Resim: 8.5. Edinilmiş tercih ve yatkınlık çalışmasında kontrol tavuklarının tedavi şeması (Johnson ve diğerleri, 1985). Işığa (A) maruz bırakılan civcivler yaklaşık 23 saatlik olduklarında, ışıklı bir odada dönen tekerleklere tek tek yerleştirildiler. Toplam maruz kalma süresi 2 saatti ve beyaz dikdörtgenlerle gösterilen iki periyottan oluşuyordu. Bu süreler arasında civcivler karanlık bir inkübatörde 1.5 saat geçirdiler. İkinci dönemden 2 veya 24 saat sonra civcivlere bir tercih testi (sırasıyla test 1 veya 2) verildi. Diğer civciv grubu (B) ise teste kadar kuvözde kaldı. Orada, bir tepe lambası (P) ile 30 dakikalık aydınlatma dışında karanlıkta tutuldular.
Dönen çarkta geçirilen süreden 2 saat sonra, "aydınlık" ve "karanlık" gruplarındaki tavukların bir kısmı tercih testine (test 1) tabi tutuldu. Her iki grubun geri kalan kuşlarına 24 saat sonra test verildi (test 2). "Koyu" gruptaki tavuklar, her iki testte de kutuyu veya doldurulmuş hayvanı tercih etmemiştir (Şekil 8.6). Test 1'deki "hafif" gruptaki tavuklar bu şekilde davrandı

Resim: 8.6. Açık ve koyu renkli tavuklar için ortalama tercihler . Hafif grup (A) civcivler dönen çarklardan kuvöze alındıktan 2 saat sonra Test 1 ve 24 saat sonra Test 1 uygulandı . "Karanlık" grup (B) aşağı yukarı aynı yaştaydı. Tercih göstergeleri formül kullanılarak hesaplandı
doldurulmuş tavuğa yaklaşmaya çalıştığında platformun kat ettiği mesafe )
Platform tarafından kat edilen toplam mesafe
100, tüm yaklaşma tepkilerinin korkuluğa yönlendirildiği anlamına gelir, 0, hepsinin kutuya yönlendirildiği anlamına gelir. (Johnson ve diğerleri, 1985.)
100
Resim: 8.7. Edinilmiş tercih ile gelişen yatkınlık arasındaki etkileşimin bir modeli . Tercih göstergeleri, Şekil 2'de sunulan verilerden ödünç alınmıştır . 8.4, C, D ve 8.6, A. Tüm ortalama tercihler, doldurulmuş tavuk tercihine göre ifade edilmiştir. Kesikli çizgiler , hafif grup tavukların karşılık gelen alt gruplarının elde ettiği test 1 ve 2'deki temel seviyeleri işaretler . AC, test 1 ve 2'deki "hafif" gruptaki tavukların ortalama tercihleri arasındaki farktır. k x -k 4 , kırmızı kutunun veya doldurulmuş tavuğun ilk sunumunun etkisini gösterir. Daha ayrıntılı açıklamalar için metne bakın. (Johnson ve diğerleri, 1985.)
aynı şekilde. Bununla birlikte, 2. testte, bu kuşlar , testten önce görülmemesine rağmen, doldurulmuş tavuğu açıkça tercih ettiler.
Tercihi tahmin etmenin olağan yöntemi, bu göstergeyi 50'ye eşit bir sıfır seviyesiyle karşılaştırmaktır. Tercihi tahmin etmenin başka bir yöntemi , "hafif" grup göstergelerini sıfır seviyesi olarak kullanmak mümkündür. Bu ölçümleri kullanırken, doldurulmuş tavuk tercihine yönelik bir önyargı dikkate alınır. Bu, Şek. 8.7. Şek . 8.4, C, D, Şek. 8.7. Bu şekilde, tüm grupların puanları - "hafif" kontrol ve ayrıca kutu ve doldurulmuş tavukla eğitilen deney grupları - doldurulmuş tavuk için tercih puanları olarak ifade edilir . Sıfır seviyesinin kaymasının değeri DE ile gösterilir. Deneysel ve "hafif" grupların ortalama tercihleri arasındaki fark k\ ile gösterilmektedir.Bu fark tavuğun edimsel öğrenme sürecinde ürettiği nesnelerden birini tercih etmesinin bir göstergesidir.
Test 1'de kutu eğitimli civcivlerin ortalama tercih değerleri "hafif" grubunkinden %19,2 daha düşüktü (Şekil 8.7, k\). Heykelle eğitilen civcivler için karşılık gelen fark, büyüklük olarak neredeyse aynıydı , yani %20,7'ydi (Şekil 8.7 ve 2 ), ancak bu şekilde kullanılan ölçeğin tersiydi . Her üç grupta da doldurulmuş tavuk tercihi 1. testten 2. teste doğru artıyor. Bu kayma, kazanılan tercihin kaybolmasından mı kaynaklanıyor, yoksa bu tercih korunuyor mu? Eğer kaybolursa, k 3 <kı ve k4<kz olması beklenmelidir (Şekil 8.7) . Bu varsayımı test etmek için, her bir testteki kontrol ışık grubunun ortalama tercih puanı , aynı testteki her deneysel kuşun tercih puanından çıkarıldı . Ortaya çıkan göstergeler istatistiksel analize tabi tutuldu. Deney kuşlarında, bu göstergeler önemli ölçüde test zamanına bağlı değildi (test 1 veya 2). Bu sonuç, edinilen tercihin çalışma süresi boyunca istikrarlı olduğunu göstermektedir. Doldurulmuş tavuğun göreceli tercihinin test 1'den test 2'ye arttığı verileriyle birlikte , bu gerçek iki sürecin deney tavuklarının tercihi üzerinde etkili olduğunu göstermektedir: tavuk grupları; 2) civcivlerin kendilerine gösterilen belirli nesneleri tanımaya başladıkları öğrenme süreci . Cisim tavuk başı ise yatkınlık artar, kırmızı kutu ise yatkınlık maskelenir ama ortadan kaldırılmaz.
1981) ördek yavrularının kendi türlerine yönelik tercihlerinin geometrik şekiller veya diğer su kuşu türleri gösterilerek giderilebileceği veya kısmen ortadan kaldırılabileceği verileriyle tutarlıdır . Ayrıca, görsel güdümlü davranışın gelişiminin ve bunun altında yatan bazı sinirsel süreçlerin hem olgunlaşma hem de deneyim tarafından değiştirildiğine dair diğer kanıtlarla da tutarlıdır (bkz. örneğin, Klopfer, 1967; Gottlieb, 1976; Bateson, 1978; Wiesel, 1982). ; Kovach, 1985).
Yatkınlığın tezahürü için koşullar
Tavukların "hafif" kontrol grubuna ilişkin veriler , deneyimlerinin bazı yönlerinin test 2'deki doldurulmuş tavuğun tercih edilmesine katkıda bulunduğunu göstermektedir. Peki hangileri? Her halükarda, "karanlık" grubun deneyiminde değillerdi, çünkü bu grup testlerin hiçbirinde tercih göstermedi. "Karanlık" ve "aydınlık" gruplar en az iki açıdan farklılık gösteriyordu. "Hafif" grup 2 saat boyunca görsel bir deneyim yaşama fırsatı buldu; ayrıca çıkrık içinde 2 saat geçirdi. Test 2'de gösterilen doldurulmuş tavuk tercihi için bu farklılıklardan hangisi anlamlıydı? Civcivler ışıkta karanlıkta olduğundan daha fazla hareket ettiğinden ( Bolhuis ve diğerleri, 1985), bu nedenle ışığa maruz kalmanın ve hareketin etkileri etkileşime girer. Ancak sadece aydınlatmayı kaldırmak ve civcivleri 2 saat dönen bir çarkta tutmanın etkisini araştırmak yeterlidir.
Bu faktörleri incelemek için üç grup tavuk alındı (Bolhuis ve diğerleri, 1985). Bir grubun civcivleri (“hafif başlangıçlı karanlık”) , inkübatörün 30 dakika dışında karanlık bir notta olan ayrı bölmelerinde kaldı (Şekil 8.8, E). Bu grup için deney koşulları, Johnson ve diğerlerinin (Johnson ve diğerleri, 1985) (Şekil 8.5, B) deneyimindeki ile aynıydı . Diğer grup ("ışıkla tetiklenen topaç"), tamamen karanlıkta bir çıkrık içinde 2 saat geçirmeleri dışında aynı koşullar altındaydı (Şekil 8.8b ). Birinci ve ikinci gruptaki tavukların davranışlarındaki herhangi bir farklılık, çıkrık içinde olma deneyimine bağlı olmalıdır. Ek olarak üçüncü grup kullanıldı (Şekil 8.8, B). Bu gruptaki civcivler (“dark spinning ”) sürekli karanlıkta, yani 30 dakikalık ışık periyodu olmaksızın tutulmuş ve çıkrıkta geçirdikleri süre dışında sürekli kuluçka makinesinde bekletilmiştir.
Önceki çalışmada olduğu gibi (Şekil 8.6, C), kuluçka makinesinde kalan koyu renkli grup civcivlerde herhangi bir nesne tercihi gelişmedi (Şekil 8.9, E). tavuklar, tarafından
teet 1 Test 2 0 20 30 52
24 .
6' p—
8' H- ^
Resim: 8.8. Doldurulmuş tavuk tercihinin ortaya çıkmasına katkıda bulunan bazı faktörleri incelemek için bir deney tasarımı (Bolhuis ve diğerleri, 1985). A ve B gruplarındaki civcivler 30 dakika (P) gün ışığına maruz bırakıldı. A grubundaki ("karanlık") civcivler, zamanın geri kalanını karanlık bir inkübatörün izole edilmiş bölmelerinde geçirdiler. Grup B'deki ("döner ") civcivler aynı koşullarda tutuldu ancak karanlıkta 1'er saatlik iki periyot halinde dönen tekerleklere yerleştirildi. Bu dönemler siyah dikdörtgenlerle işaretlenmiştir; aralarında 1.5 saat ara verildi Grup B civcivler , karanlık çıkrıkta geçirilen 1'er saatlik iki dönem dışında her zaman karanlık inkübatörde sosyal olarak izole kaldılar . Tüm civcivlerin tercihi 30 veya 52 saatlik olduklarında değerlendirilmiştir.
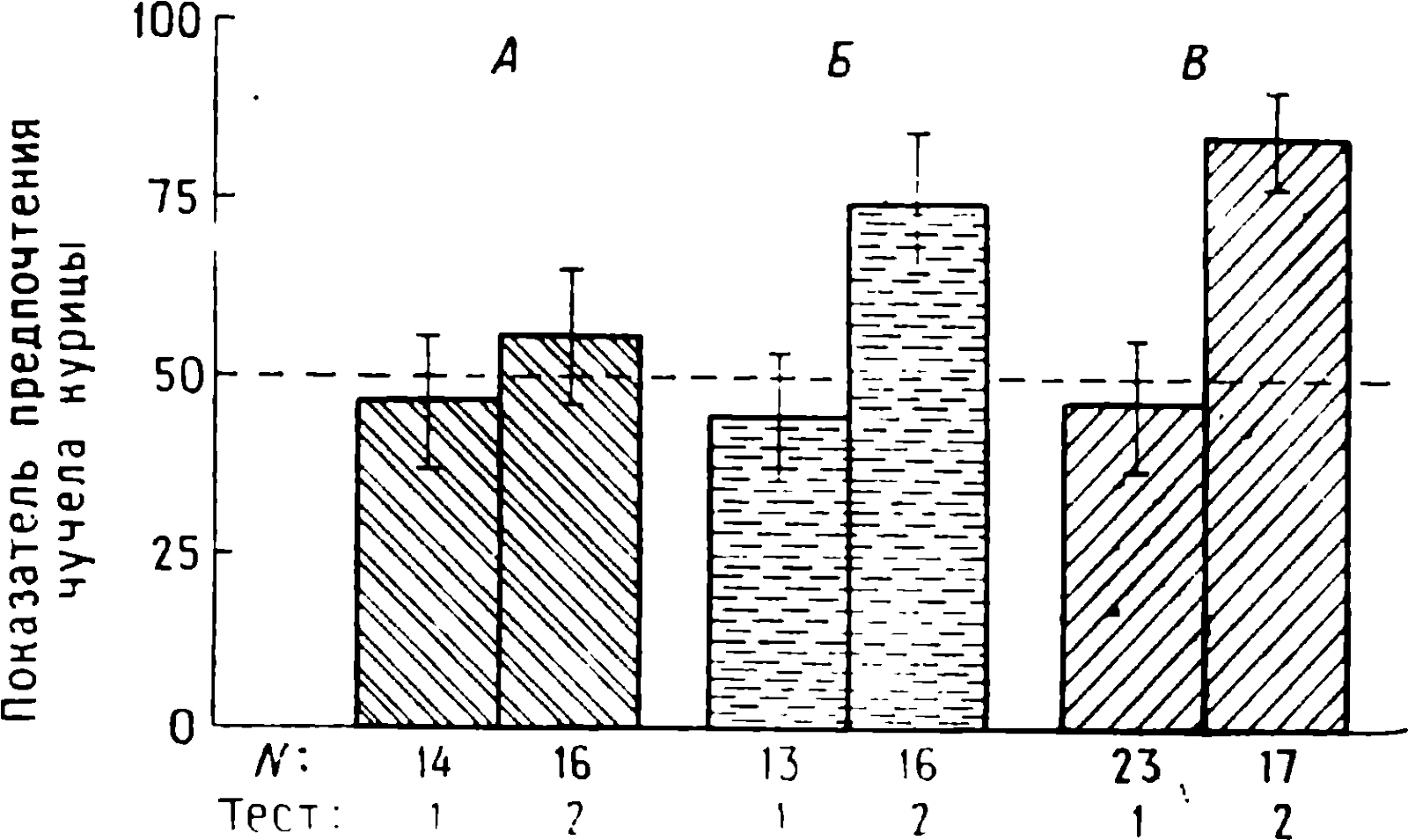
Resim: 8.9. Şekil 1'de belirtilen koşullar altında tavuklarda tercih göstergeleri. Sırasıyla 8.8, A, B ve C. Göstergeler, şekil 2'nin başlığında verilen formüle göre hesaplanmıştır. 8.6, A. Başlangıçta grup A'daki civcivler ışığa maruz bırakıldı ve geri kalan süre tercihlerini test etmeden önce karanlık bir inkübatörde kaldı. B grubundaki tavuklar önce ışığa maruz bırakılmış, ardından dönen bir çarkta 2 saat geçirilmiştir. Grup B civcivler seçim testinden önce her zaman karanlıkta tutuldu; Karanlık bir çıkrık içinde 2 saat geçirdiler. (Bolhuis ve diğerleri, 1985.)
30 dakikalık ön aydınlatma alan ve ardından 2 saati karanlık bir çıkrık içinde geçirenler test 2'de doldurulmuş tavuğu tercih ederken test 1'de tercih etmediler (Şekil 8.9, B). Son olarak, her zaman karanlıkta tutulan ancak çıkrık içinde 2 saat geçiren civcivler de test 2'de doldurulmuş tavuğu tercih ettiklerini gösterdiler (Şekil 8.9, B). Test 2'deki çarktaki her bir tavuk grubunun aktivitesi, test 1'deki aktiviteden önemli ölçüde farklı değildi.
Bu deneyin sonuçları, çıkrık içinde olma deneyiminin ve/veya tavuklarla uğraşmak gibi ilgili bazı deneyimlerin 24 saat sonra bir tercih oluşturmak için yeterli olduğunu göstermektedir . görsel deneyimin
doldurulmuş tavuk tercihinin tezahürünü etkilemediğini varsaymak yanlış olur . Böyle bir etki gerçekten de iki grup tavuk üzerinde yapılan bir deneyde gösterildi (Bolhuis ve diğerleri, 1985). Bir grubun koşulları, Johnson ve arkadaşlarının (Johnson ve diğerleri, 1985) (Şekil 8.5, 4) deneylerindeki "hafif" grup ile aynıydı. Diğer grup için koşullar biraz farklıydı. Çıkrık içindeyken, her civciv karmaşık bir geometrik desen gördü (Şekil 8.10). Önceki seride olduğu gibi (Şekil 8.6, A), "hafif" gruptaki tavuklar

zcm
Resim: 8.10. Bir grup tavuğun eğitildiği bir odanın iki duvarını kaplayan karmaşık bir desen . Desen, çıkrıktayken her tavuk tarafından görülebiliyordu. (Bolhuis ve diğerleri, 1985.)
girdikten 24 saat sonra doldurulmuş tavuğu tercih ettiğini gösterdi , ancak önceki testte göstermedi (Şekil 8.11, K). Bunun aksine, karmaşık bir geometrik desen gören tavuklar, çarka girdikten 2 saat sonra doldurulmuş bir hayvanı tercih ettiler (Şekil 8.11, B). Bu iki gruptaki farklı tercih tezahürü, lokomotor aktivitedeki farklılıklarla ilişkili değildi, çünkü her ikisi de tekerlekte ve testler sırasında eşit derecede aktifti. Bu nedenle, bu deneylerde elde edilen veriler, karmaşık kalıpları gözlemleme deneyiminin, doldurulmuş tavuk tercihinin ortaya çıkmasını hızlandırabileceğini göstermektedir. Belli ki, spesifik olmayan görsel deneyimin yanı sıra çıkrık içinde olmakla ilgili spesifik olmayan deneyim , bir kutu yerine doldurulmuş bir hayvan için belirli bir tercihin gelişimini etkileyebilir . Civcivler, testten önce doldurulmuş hayvanı görmedikleri için, onun hakkında hiçbir şey öğrenmeleri mümkün değildi. Doldurulmuş tavuğun ilk maruz kalması tavukların onu tercih etmesi için gerekli değildir.
Bu bölümde ve Sec. 8.2.1, belirli koşullar altında civcivlerin doldurulmuş tavuğa yatkın olabileceğini öne sürmektedir. Ancak bu yatkınlık, Tablo 1'de listelenen deneylerde öğrenmenin etkilerini dengelemedi. 8.1. Bu çalışmalarda tercih sıralı bir testle ölçülmüştür. Daha önce (s. 180) dolma tavuk için ortaya çıkan tercihin tespit edilememesinin , testin yetersiz hassasiyetinin bir sonucu olabileceğini söylemiştik (s. 180). Veri

Resim: 8.11. Işığa maruz kalan iki civciv grubu tercih edilir. Göstergeler, şekil 2'nin başlığında verilen formüle göre hesaplanmıştır. 8.6. A Grubu civcivler , Şekil 2'deki "hafif" grupla aynı şekilde tedavi edildi. 8.5, A. Test 1'de herhangi bir tercih göstermediler (50'ye eşit göstergeye göre), ancak test 2'de doldurulmuş tavuk tercihi anlamlıydı (P<0.01). Grup B civcivleri , grup A ile aynı koşullardaydı ancak çıkrıktayken karmaşık bir model gördüler (Şekil 8.10) . Bu civcivler her iki testte de doldurulmuş tavuğu tercih etmişlerdir (P<0.01). (Bolhuis ve diğerleri, 1985.)
Durumun gerçekten böyle olduğuna dair kanıt , karanlıkta yetiştirilen tavukların 2 saat boyunca dönen bir tekerleğe yerleştirildiği, her seferinde civcivlerin bir yarısına ardışık test ve diğer yarısına eş zamanlı test verildiği yayınlanmamış bir deneyden geliyor. Seri test verilen civcivler, her iki testte de kutuyu veya kuklayı tercih etmemiştir. Buna karşılık , eş zamanlı seleksiyon testine tabi tutulan civcivler, çıkrığa maruz kaldıktan 24 saat sonra beklenen yem tercihini sergiledi. Bu sonuçlar , saptanabilir kukla tercihinin bir seri testte ortaya çıkamayacak kadar zayıf olduğunu göstermektedir. Ancak eş zamanlı seleksiyon testinde tespit edilebilecek kadar güçlüdür ve bu nedenle civcivlerin davranışını önemli ölçüde değiştirecek kadar güçlü olabilir (s. 195).
8.3. Neye yatkınlık?
Doldurulmuş tavuğun artan tercihi, aynı türün bireylerinin bazı özelliklerine artan bir ilgiyi mi yoksa basit olanlara kıyasla karmaşık biçimlere artan bir ilgiyi mi ifade ediyor? Açıklanan deneylerin hiçbiri


yukarıdaki nyh bu soruyu cevaplamaya izin vermiyor; bu nedenle, yeni bir dizi deney düzenlendi (Johnson ve Horn, 1986a). Doldurulmuş tavuk iki boyutlu olsaydı, bu deneyleri kurmak kolay olurdu: o zaman görüntü, görsel olarak karmaşık kalırken artık bir tavuğa benzemeyecek şekilde kesilebilirdi . Civcivler kendi türlerinin üyelerini tercih ederse, böyle bir "karma" görüntü onlar için normal olandan daha az çekici olacaktır. Ancak korkuluk üç boyutlu bir nesnedir, bu nedenle soruna farklı bir yaklaşım gerekir; Kuşların derilerini elde etmek kolay olmadığı için ekonomiye de dikkat edilmelidir.
Karanlıkta tutulan civcivlerin 2 saat boyunca bir çıkrığa yerleştirildiği aynı basit davranış inceleme yöntemini kullandık. Deneyim planı, Şek. 8.8, B. Bu deneyde, seçim testinde kullanılan nesneler kırmızı bir kutu ve doldurulmuş tavuktu. Yeni deney serisinde doldurulmuş tavuk da kullanıldı, ancak kırmızı kutunun yerini Şekil 1'de gösterilen nesnelerden biri aldı. 8.12. Karanlıkta çıkrık içinde geçirilen sürenin bitiminden 2 veya 24 saat sonra tercih sınandı . Doldurulmuş tavukla eşleştirildiğinde , kırmızı bir kutu sunuldu, doldurulmuş tavuk tercihi test 2'de göründü, ancak test 1'de görülmedi (Şekil 8.9, B). Yeni alternatif nesneler, tüm doldurulmuş hayvanla aynı yanıtı ortaya çıkarsa, tercih puanları her iki testte de 50'den önemli ölçüde farklı olmayacaktır.
Şek. 8.12, A, çeşitli parçaları doğal olmayan bir konumda birbirine tutturulmuş bir tavuk kaşıydı . Bununla birlikte, bir insan için, bu nesne şüphe götürmez bir şekilde bir tavuğa benziyordu. Görünüşe göre bu tavuklar için de geçerliydi. Her iki durumda da normal doldurulmuş hayvanı modifikasyonuna tercih etmediler.
100
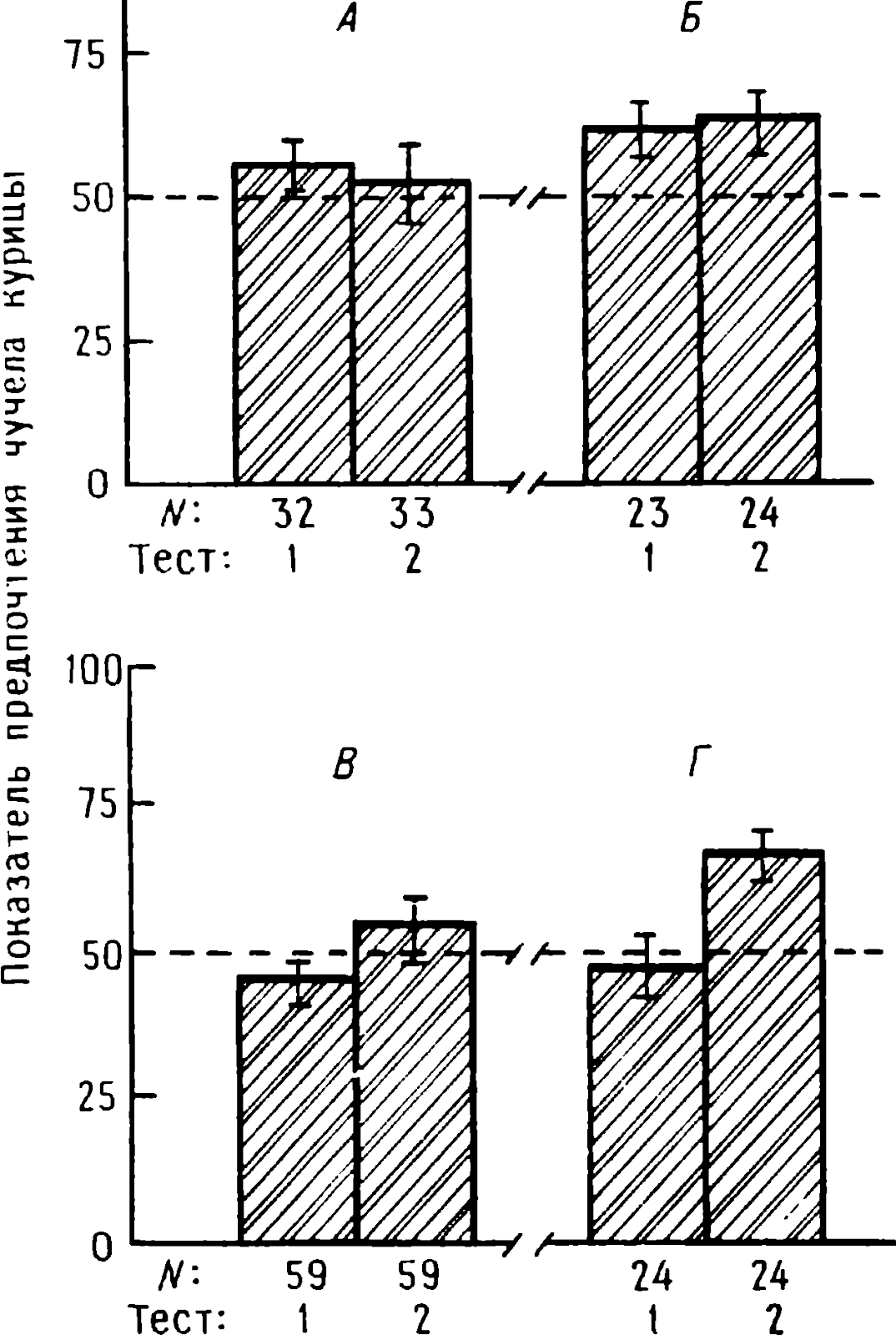
Resim: 8.13. Doldurulmuş tavuk ile Şekil 1'de gösterilen nesnelerden biri arasında seçim yapan tavuklarda tercih indeksleri. 8.12 ve karşılık gelen harflerle işaretlenmiştir. Deneysel tasarım, Şekil l'deki ile aynıydı. 8.8 B. Test 1 civcivler dönen çarka girdikten 2 saat sonra ve Test 2 24 saat sonra yapıldı. Şekil 1'de gösterilen nesne için yalnızca test 2'deki gösterge. 8.12, D, test 1'dekinden önemli ölçüde yüksekti (P<0.001). (Johnson, Bacaklar, 1986a.)
testler (Şekil 8.13, D). Alternatif nesne kısmen parçalanmış bir doldurulmuş tavuk olduğunda sonuçlar çok benzerdi (Şekil 8.12, B). Doldurulmuş hayvanın tamamı için hafif bir tercih vardı ve her iki testteki puanlar hemen hemen aynıydı (Şekil 8.13, B). Şek. 8.12, B, doldurulmuş bir hayvanın büyük parçalarından oluşuyordu ve 1. ve 2. testlerin sonuçları yine birbirine yakın çıktı (Şekil 8.13, B). Son olarak, kuşun derisi parçalara ayrıldı ve kutunun iki geniş duvarına sabitlendi (Şekil 8.12, D). Test 1'de civcivler herhangi bir tercih göstermedi, ancak test 2'de açıkça bütün doldurulmuş tavuğu tercih ettiler (Şekil 8.13, D).
Ortaya çıkan tercih (Şekil 8.13d ) , Şekil 1'de gösterilen nesnelerin karmaşıklığından dolayı mankenin yüzeyinin daha yüksek karmaşıklığına bağlı görünmüyordu . 8.12, D, C, Şek. 8.12, D. Test 2'deki tavukların bir tavuğun karmaşık dış hatlarını bir kutunun dış hatlarına tercih etmesi de olası değildir: Şekil 2'de gösterilen nesne yapılırken doldurulmuş parçalar kutuya iliştirilmiştir. 8.12B, ancak test 2'deki tavuklar için bütün bir doldurulmuş tavuk kadar çekiciydi (Şekil 8.13B).
Bu deneylerin sonuçları, doldurulmuş bir tavuğun tercih edilmesinin, nesnenin doldurulmuş olandan karmaşıklık seviyesindeki farkla açıklanmasının zor olduğunu göstermektedir . Gelişen tercihin, türünün temsilcisi olarak tanımlanabilecek yeterli sayıda öğeye sahip nesnelere yönelik olduğunu iddia etmek mümkün hale gelir . Bu olasılık, civcivin doldurulmuş bir tavuk ile başka türden doldurulmuş bir kuş (Johnson ve Hoth, 1986a), yani gri ördek (Anas strepera) arasında seçim yapmasına izin verilerek test edilmiştir . Deneyin planı öncekiyle aynıydı. Civcivler her iki testte de tavuğu veya ördeği tercih etmemiştir. Doldurulmuş bir tavuk ile bir memeli - gelincik (Mustela putorius) arasında seçim yaparken aynı sonuçlar elde edildi.
Bu deneyler, gösterilen tercihin özellikle aynı türden veya cinsten hayvanlara yönelik olduğunu doğrulamamıştır. Bunun yerine, tercihin tamamen ve kısmen dönüştürülmüş doldurulmuş hayvanlarda bulunan bazı öğelere yönelik olduğunu gösterirler (Şekil 8.12, E-C), ancak Şekil 1'de gösterilen kırmızı kutuda veya deri parçalarının yapısında yoktur. 8.12, D. Bu öğelerin ne olduğu bilinmiyor, ancak nispeten basit (Mugg ve Nishihara, 1978) veya baş ve gözler gibi çok spesifik (Tinbergen ve Perdeck, 1950; Tinbergen, 1951; Hinde, 1954; Coss) olabilirler. , 1972).
Doldurulmuş tavuğun tercih edilmesinde PMVG'nin rolü nedir?
Tercih geliştirme
Sağlam tavuklar üzerinde kuş tercihinin gelişimini ortaya koyan deneyler yapıldı. PMVP bu tercihin gelişimine katılıyor mu veya PMGP'nin yok olmasına rağmen kendini gösteriyor mu, değişen tercih PMVP'nin dışında kalan farazi tanıma sistemindeki süreçlerin bir sonucuysa beklenebileceği gibi (s. 179) )?
bir cismi aydınlatmak ve döndürmek için pedala basmayı öğrendiğinde , bu pekiştirici cismin özelliklerini öğrenir (Bölüm 6.2). PMVG'si zarar görmüş civcivler bu edimsel yanıtı öğrenebilir (bkz. sayfa 131 ve şekil 6.4). Ancak, tercihleri 2 saat sonra test edildiğinde nesne hakkında çok az şey biliyor gibi görünüyorlar (bkz. Şekil 6.5). Şek. Şekil 6.5, kutu ve doldurulmuş tavukla eğitilmiş civcivlerin puanlarını birleştirir. Edimsel öğrenmede kullanılan pekiştirici nesneye göre bölünmüş aynı sonuçlar, Şekil 1'de gösterilmektedir. 8.14, A. PMVP hasar gördüğünde takviye nesnesinin tercihi ihlal edildi. Bu testin sonunda civcivler tekrar karanlık inkübatöre alındı. Tercih, eğitimin bitiminden 24 saat sonra tekrar test edildi (Şekil 8.14, B). Sahte ameliyat edilen tavukların puanları, eğitimde hangi nesnenin kullanıldığına bakılmaksızın test 1'den test 2'ye önemli ölçüde doldurulmuş tavuk lehine değişti. PMVG'si zarar görmüş kuşlarda da benzer bir değişim meydana geldi. Bu nedenle, ortaya çıkan tercih, PMVG'nin korunmasına bağlı değildir.
Bireysel Tanıma
PMVP'nin bireysel tanımaya katılma olasılığına ilişkin çalışma, üç grup tavuk üzerinde gerçekleştirildi (Johnson ve Horn, 1986c, 1987). 16 civcive çıkıştan kısa bir süre sonra bilateral Wulst yıkımı , diğer 16 civcivde ise PMVG uygulandı. Sahte ameliyatlı kontrol grubu 52 civcivden oluştu. Ertesi gün, civcivler yaklaşık 30 saatlik olduklarında, dönen bir tekerleğe tek tek yerleştirildiler ve 200 dakika boyunca doldurulmuş tavuk verildi. İki doldurulmuş hayvan kullanılmış ve tavukların yarısına bunlardan biri, diğer yarısına ikincisi takdim edilmiştir (Şekil 8.15). Tercih, her iki mankenin de uçlarına yerleştirildiği eş zamanlı bir seçim aparatında ölçülmüştür. Test, eğitimden 3 saatten daha kısa bir süre sonra gerçekleştirildi. Sahte çalışan grubu kontrol edin
13-140
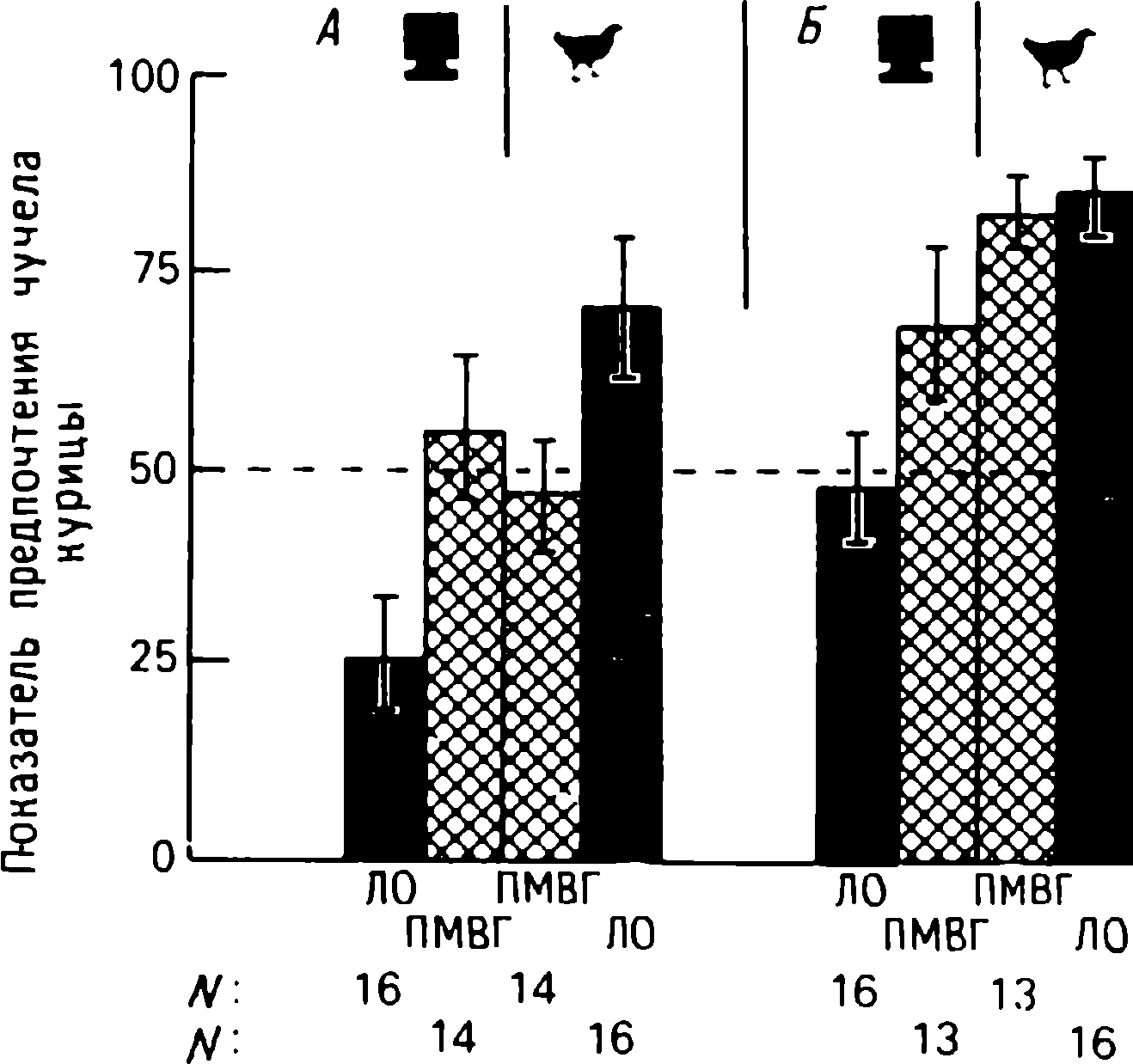
Resim: 8.14. İlişkisel öğrenme sırasında bir kutuya veya doldurulmuş tavuğa maruz kalan civcivlerde tercih puanları (Bölüm 6.2). Tercih yanıtı , eğitimden 2 saat (A) ve 24 saat (B) sonra test edildi . Eğitimden önce PMVG'nin iki taraflı imhası yapıldı. Tercih göstergeleri, Şekil 2'nin başlığında verilen formül kullanılarak hesaplanmıştır. 8.6; tavuk dolması tercihini yansıttılar. 50'nin oldukça altındaki bir puan , kırmızı kutu için bir tercihi yansıtır ve 50'nin üzerindeki bir puan, korkuluk için bir tercihi gösterir. Eğitimden 2 saat sonra test edildiğinde (Test 1), sahte ameliyat edilen tavuk grupları daha önce gördükleri nesneyi tercih ettiler (P<0.05). Daha önce kutuyu görmüş olan hasarlı PMWH'li civcivlerin puanları , kontrol grubundan önemli ölçüde farklıydı (P < 0.02). Doldurulmuş tavuğu gören tavukların indekslerindeki fark anlamlılık eşiğindeydi (/= 1.91; 0.1<P>0.05). Test 2'deki tercih yeniden değerlendirilirken, tüm gruplar doldurulmuş tavuk tercihinde önemli bir artış gösterdi (tüm durumlarda P < 0.01). (Johnson, Noth, 1986b.)
baba ve Wulst'u zarar görmüş civcivler tanıdık doldurulmuş tavuğu tercih ettiler; ortalama tercih puanları (sırasıyla 63.6 ve 69.7), rasgele 50 seviyesinden (P <0.05) anlamlı derecede yüksekti . Buna karşılık, PMA'sı hasar görmüş civcivlerin ortalama puanı (38.3) 50'den önemli ölçüde farklı değildi. Ortalama puanları diğer iki grubunkinden önemli ölçüde düşüktü (her durumda P < 0.02). Deneyin sonuçları , zarar görmemiş tavukların yanı sıra zarar görmüş Wulst'a sahip tavukların , gözlemlenen kuşun bazı özelliklerini hatırladıklarını ve daha sonra bu bireyi tanıyabildiklerini göstermektedir. Görünüşe göre PMVP bu tanıma işlemi için gerekli.
Bu bölümde ve ayrıca Bölüm'de sunulan veriler. 8.2, 8.3 ve 8.4.1 , PMGB'nin dışında kalan ve belirli morfolojik özelliklere sahip nesnelere doğuştan gelen yaklaşma arzusunu destekleyen özel bir nöron sisteminin varlığı hipoteziyle uyumludur . Bu

Resim: 8.15. Bireysel tanımada PMVG'nin rolüne ilişkin çalışmada kullanılan iki doldurulmuş Banking tavuğunun görüntüsü.
önerilen sistem açıkça yumurtadan çıktıktan hemen sonra çalışmaz , ancak çeşitli etkilerle tetiklenebilir , örneğin, bir civcivi 2 saat boyunca dönen bir tekerleğe yerleştirmek veya ona karmaşık bir görsel kompleks sunmak. Klopfer ( 1967) ayrıca ördek yavrularında yatkınlığı tetiklemek için aktivasyon ihtiyacını bulmuştur. Şöyle yazıyor: " Takip etme sürecinde etkinleştirilen bir tür doğuştan tercih var gibi görünüyor." Bunun altında yatan fizyolojik süreçler ve spesifik olmayan deneyimlerin onları nasıl tetiklediği daha sonra tartışılacaktır (Bölüm 9.6, 10.3.7 ve 12.2.2).
Ortaya çıkan tercih, bir civcivin erken yaşamında önemli bir rol oynayabilir. Tavukların, ortamın görsel karmaşıklığına tek bir kez maruz kalmayla veya ortamdaki hareketin bir sonucu olarak (karanlıkta bile), uyaranın belirli özelliklerini, örneğin kafalarını veya gözlerini hızlı bir şekilde tercih etmesi mümkündür. Önemli olmak. Diğer hayvanlar da bu elementlere sahiptir , ancak yumurtadan çıktıktan hemen sonra anneyi görme olasılığı en yüksektir. İlk tercih çok hafif olabilir (s. 188-189), ancak civcivlerin kendi türlerine yaklaşmasına neden olacak kadar yeterlidir. Öğrenme süreci daha sonra bu tercihi bir kişiyle sınırlayabilir. Böylece ilk tercih, civcivi büyük olasılıkla aynı türden bir yetişkin olan uygun nesneye yönlendirecektir . Bu durumda öğrenme süreci, kişinin kendi annesinin bireysel özelliklerini tanımasının ek bir yolu olarak görülebilir. Bu nedenle, yaşamın ilk saatlerinde ve günlerinde, spesifik olmayan deneyimler , civcivin kendi türünü tanımasını sağlayan sistemi harekete geçirebilir; özel deneyim, 13*
öğrenme sürecinde geliştirilen, tavuğun akrabalarını tanımasını sağlar. Görünüşe göre bu öğrenme sürecinde PMWG yer alıyor.
Depolama sistemi X' ve doldurulmuş tavuk
bölümde alıntılanmıştır. Şekil 7'deki veriler, PMVP'nin dışında bulunan ikinci depolamanın ( Xx ), baskı sırasında kademeli olarak oluştuğunu göstermektedir. Doldurulmuş tavuk tercihini sürdürebilen bir tanıma sistemi SMP'nin dışında da bulunduğundan (Bölüm 8.1 ve 8.4.1), bu sistemlerin aynı olup olmadığını bilmek gerekir. Bağımsız olduklarına inanmak için sebepler var.
Tabloda listelenen deneylerde. 8.1, PMVG'ye verilen hasar, yalnızca kutu eğitimli civcivlerde tercih puanlarını düşürdü. PMCH yıkımının tercih yanıtı üzerinde hiçbir etkisinin olmadığı durumlarda (Bölüm 7.1 ve Şekil 7.3, lPMCH grubu), sapmalar yalnızca kutunun sunumu ile eğitilen grupta gözlendi. WWGP dışında X' deposunun varlığını varsaymanın ilk nedeni , bu kuşların davranışını açıklama ihtiyacıydı. Bu varsayım, eğitimden 26 saat sonra PMIG'deki iki taraflı hasarın tercihleri önemli ölçüde etkilemediği gerçeğiyle desteklenmektedir (Şekil 7.6, B). Şekil 2'de sunulan verilerin müteakip analizi. 7.6, B, kutu ve doldurulmuş tavuk sunumu ile eğitilen gruplar için PMSH'si zarar görmüş tavuklarda ortalama tercih değerlerinin sırasıyla 74.6±6.8 ve 81.4±8.2 olduğunu gösterdi. Her iki ortalama da anlamlı olarak 50'den yüksektir (P<0.005). Bu nedenle X' , tavuğun tanımayı öğrendiği nesne hakkında bilgi depolayabilir; X'in , kendi türünün üyeleri de dahil olmak üzere belirli nesnelere yaklaşma eğilimini koruyan PMHG dışında da varsayılan sistemle herhangi bir ilgisi olduğuna inanmak için hiçbir neden yoktur .
Bu düşünceler, X ve X' sistemlerinin genel özellikleri fikrini büyük ölçüde basitleştirir (Bölüm 7.3 ve Şekil 7.8, K): öğrenme sürecinde elde edilen bilgilerin depolanmasına katılırlar .
8.6 Özet
PMSH'ye verilen hasarın, tanıdık bir uyaran tercihini bozduğu deneylerde, öğrenme sırasında hangi uyaranın sunulduğuna bakılmaksızın bu gözlemlendi. Bununla birlikte, kutu eğitimli civcivlerde azalma önemliydi ve doldurulmuş tavuk eğitimli grupta nispeten zayıftı. Bu sonuçlar, kendi türünden bireyleri tanıma bağlamında tartışılmıştır . Tercihi değerlendirmek için hassas bir test kullanıldığında, doldurulmuş bir tavuğa göre yaşamın ilk 2-3 gününde arttığı gösterilmiştir. Bu tercih, eğitimde hangi nesne kullanılırsa kullanılsın kendini gösterir. Görsel deneyime sahip olan ancak tercih gücü ölçülene kadar hiçbir nesneyi görmemiş tavuklarda kendini gösterir . Ayrıca karanlıkta tutulan civcivler bile 2 saat çıkrık içinde kalmayı tercih ediyor. Her zaman karanlık bir inkübatörün ayrı bölmelerinde olan civcivlerde görülmez . Tercihin iki sürece dayandığı sonucu çıkar. İlk olarak, doldurulmuş tavuğun çekiciliğinde bir artış olarak kendini gösteren gelişen bir yatkınlık vardır. Bu yatkınlık, spesifik olmayan deneyimler tarafından tetiklenebilir ve ortaya çıktıktan sonra öğrenme süreciyle etkileşime girebilir. İkincisi, civcivlerin belirli nesneleri tanıma yeteneğini kazandığı öğrenme sürecidir. PMVP, ikinci süreçle ilgilidir. Bilateral MVH hasarı olan civcivlerde büst tercihi ortaya çıktığı için PMSH dışındaki bazı yapılar ilk işlemi sağlar. Ortaya çıkan tercihin, basit olanlara kıyasla karmaşık görsel nesnelere veya yalnızca aynı türün temsilcilerinde bulunan morfolojik özelliklere bir yatkınlığı yansıtmadığı gösterilmiştir .
EDEBİYAT
Bateson R. R. G. Eryu deneyimi ve cinsel tercihleri. İçinde: Cinsel davranışın biyolojik belirleyicileri {ed. JB Hutchinson, 1978, s. 29-53. Chichester'dan John Wiley.
Bolhuis Johnson MH, Horn G. Evcil civciv Devi'de evlada tercihlerinin gelişimi üzerinde eury deneyiminin etkileri. Psychobiol., 1985, 18, 299-308.
Cipolla-Neto J., Horn G., McCabe BJ Hemisferik asimetri ve damgalama: sıralı lezyonların hyperstriatum ventrale'ye etkisi, Exp. Brain Res, 1982, 48, 22-27.
Coss PG Göz benzeri şemalar: davranış üzerindeki etkileri. Doktora tezi, Reading Üniversitesi, 1972.
Fabricius E., Boyd H. Ördek yavrularının aşağıdaki tepkileri üzerine deneyler, Wildfowl Trust Ann. Rep., 1952/53, 6, 84-89.
Gottlieb G. Deneyimin davranış ve sinir sisteminin gelişimindeki rolü. In: Davranış ve sinir sisteminin gelişimi üzerine çalışmalar , Cilt. 3, Nöral ve davranışsal özgüllük (ed. G. Gottlieb), 1976, s. 25-54, Academic Press, New York.
Hindu s. A. İspinozun (Fringilla coelebs) yıldırma davranışında gösterildiği gibi, kısmen Doğuştan gelen bir tepkinin gücündeki değişiklikleri yöneten faktörler. I. Yanıtın niteliği ve seyrinin incelenmesi,
Proc. R. Soc, 1954, B142, 306-331.
Not G., McCabe B. J. Önyargılar ve tercihler. Lezyonların civciv beynine damgalanması üzerindeki etkiler, Anim. davran." 1984, 32, 288-292.
Not G., McCabe B. J., Cipolla-Neto J. Evcil civcivde damgalama: hiperstriatum ventrale'nin her iki tarafının edinme ve tutmadaki rolü, Exp. Brain Res., 1983, 53, 91-98.
Johnson MH, Horn G. Evcil civciv Behav'da bir yatkınlığın analizi. Brain Res., 1986a, 20, 108-109.
Johnson MH, Horn G. Civciv ön beyninin sınırlı bir lezyonu ile tanıma belleği ve çağrışımsal öğrenmenin ayrışması, Neuropsychologia, 1986b, 24, 329-340.
Johnson MH, Horn G. Evcil civcivlerin sınırlı bir beyin bölgesi, bireysel türdeşlerin tanınmasında yer alıyor mu? davranış. Brain Res., 1986c, 20, 109-110.
Johnson MH, Horn G, Civciv ön beyninin sınırlı bir bölgesinin, bireyin tanınmasındaki rolü, Behav. Brain Res., 1987 (baskıda).
Johnson MH, Bolhuis JJ, Horn G. Baskı sırasında edinilen tercihler ile gelişen yatkınlıklar arasındaki etkileşim. Animasyon Behav., 1985, 33, 1000-1006.
Johnston TD, Gottlieb G. Ördek yavrularında görsel türlerin tanımlanması, Anim. Behav., 1981, 29, 1082-1099.
Klopfer PH Cheshire kedisi mi damgalıyor? davranış. Sci., 1967, 12, 122-129.
Kovach JK Civcivlerde yaklaşma davranışının ortaya çıkarılması ve geliştirilmesinde farklı renklerin etkinliği, Behavior, 1971, 38, 154-168.
Kovach JK Algısal damgalama: genetik olarak değişken tepki eğilimleri, seçici öğrenme ve bıldırcın civcivlerinde (C. coturnix Japonica) renk ve desen tercihlerinin fenotipik ifadesi , Behaviour, 1983, 86, 72-88.
Kovach JK Davranış genetiği ve engram arayışı: genler, algısal tercihler ve uyarıcı Bilginin aracılığı. İçinde: Davranış genetiği: ilkeler ve uygulamalar II (ed. JL Fuller ve EC Simmel), Erlbaum, Hillsdale, NJ 1985.
McCabe BJ, Cipolla-Neto J., Horn G., Bateson PPG Baskıdan sonra civcivin hiperstriatum ventraline yerleştirilen bilateral lezyonların amnezik etkileri, Exp. Brain Res., 1982, 48, 13-21.
McCabe BJ, Horn G., Bateson PPG Civciv ön beyninin sınırlı lezyonlarının damgalama sırasında evlat tercihlerinin edinilmesi üzerindeki etkileri, Brain Res., 1981, 205, 29-37.
Mackintosh NJ The Psychology of Animal Learning, Academic Press, Londra, 1974.
Marr D., Nishihara HK Üç boyutlu şekillerin uzamsal organizasyonunun temsili ve tanınması, Proc. R. Soc., 1978, B200, 269-294.
Schaefer HH, Hess EH Baskı nesnelerinde renk tercihleri, Z. Tierpsychol, 1959, 16, 161-172.
Sluckin W. Damgalama ve erken öğrenme, Methuen, Londra, 1972.
Tinbergen N. İçgüdü çalışması. Clarendon Press, Oxford, 1951.
Tinbergen N., Perdeck AC Yumurtadan yeni çıkmış ringa martı civcivindeki yalvarma tepkisini serbest bırakan uyaran durumu hakkında. (Larus a. argentatus Pont), Davranış, 1950, 3, 1-39.
Wiesel TN Doğum sonrası görsel korteksin gelişimi ve çevrenin etkisi, Nature, Londra., 1982, 299, 583-591.
BÖLÜM 9
HAFIZA VE İZLEMENİN FİZYOLOJİK SINIRLARI
Ch'de. Şekil 6'da, baskı yapan nesne hakkındaki bilgilerin yalnızca depolama sistemi, motivasyon sisteminden gelen bir sinyalle aynı anda nesneden duyusal bir sinyal aldığında kaydedilmesi önerildi (Bölüm 6.4). Bu varsayım doğruysa, sadece duyusal bir girdi alındığında, bellek sisteminde izlerin oluşması imkansızdır. Girdinin bu şekilde sınırlandırıldığı bir deney tasarlayabildiğimizi ve ardından onu çağıran nesneyi tavuklara gösterebildiğimizi varsayalım . İzler oluşmadığı için civcivler civcivlerin civciv tercihi göstermemelidir. Benzer bir çalışma yapılmıştır (McCabe ve diğerleri, 1979). Deney, doğrudan PMVP'ye yapay olarak bilgi verme olasılığını araştıracak ve bu prosedürün tercihi etkileyip etkilemediğini keşfedecek şekilde organize edildi . Sonuç pozitif olsaydı, bu , PMVP'nin baskıya dahil edilmesi lehine başka bir kanıt olurdu.
Beyin stimülasyonunun etkileri
Tavuklar, 4,5 ve 1,5 Hz frekanslı ışık flaşlarını ayırt edebilir. Bunu gösteren deneylerde, bir silindir döndürülerek titreyen ışık üretildi (Şekil 9.1). Civcivlere bir tercih oluşturmak için bu frekanslardan birinin 5 saat boyunca sunulması gerektiği tespit edildi.Nörofizyolojik açıdan bakıldığında, titreşen ışık iki nedenden dolayı çok uygun bir uyarandır. İlk olarak, uyarılan uyaranın uygun frekansına göre uyarılmış yanıtları kaydetmek ve böylece bu "etiketli" yanıtların sinir sistemindeki yayılma yollarını ortaya çıkarmak mümkündür. İkincisi, sinir sisteminin herhangi bir bölgesini belirli bir frekanstaki dürtülerle uyarmak ve böylece hayvanın karşılık gelen frekansta bir ışık parlaması tercihi geliştirip geliştirmediğini ortaya çıkarmak kolaydır. İkinci yöntem, PMVP'nin bu şekilde uyarılmasının civcivin müteakip yaklaşma tepkilerini etkileyip etkilemediğini bulmak için kullanıldı.

Resim: 9.1. Seçim testinde kullanılan görsel uyarıcının kesit diyagramı. Silindir, 48 watt'lık bir ampulle içeriden aydınlatıldı. Işık , silindiri kaplayan aydınger kağıdından geçti. Bir silindirde bir, diğerinde üç siyah şerit yapıldı. Silindir sabit bir frekansta döndüğünde, şeritlerin sayısına bağlı olarak 1,5 veya 4,5 Hz frekansta titreyen ışık yarıktan görülebiliyordu. Mat siyah yüzeye 15x5 cm'lik bir kesi yapıldı (McCabe ve ark., 1979).
Civcivler, yumurtadan çıktıkları andan 10 saat yaşına kadar karanlıkta tutuldu, ardından anestezi uygulanarak her iki yarım küreye bipolar uyarıcı elektrotlar yerleştirildi. Elektrotlar SMVG'ye, ek hiperstriatuma ve ekstostriatuma yerleştirildi. Son iki alan kontrol olarak seçildi. Her ikisi de görsel projeksiyon bölgeleridir, bu nedenle SMVG'ye giden herhangi bir görsel sinyalin bu bölgeleri de etkilemesi gerektiği varsayılmıştır (Bölüm 11.1.1). Ameliyattan 16 saat sonra civcivler, ses geçirmez bir odada bulunan ayrı ayrı dönen tekerleklere yerleştirildi. Her tekerleğin merkezi, arkasında çizgili bir silindir bulunan mat siyah yüzeyden 50 cm uzaktaydı (Şekil 9.1). Çarkta kaldığı süre boyunca, tavuğun beyninin herhangi bir kısmı, 1 saniyede 1,5 veya 4,5 darbe frekansında bir dizi elektriksel darbe ile uyarıldı. Stimülasyon sırasında, civcivin siyah yüzeydeki bir yarıktan görebilmesi için silindirli kutu aydınlatıldı . Silindir ayrıca içeriden aydınlatıldı ve şeffaf şeritleri sarı görünüyordu. Beyin uyarılırken silindir hareketsiz kaldı. Tavuklar yerleştirildi

1 saniyedeki uyaran sayısı *1,5 4,5
Resim: 9.2. PMVG stimülasyonundan sonra civcivlerde ortalama tercih puanları. Gösterge, platformun orta noktadan 10 dakikada kat ettiği maksimum mesafe olarak alınmıştır (Bateson ve Wainwright, 1972). Y eksenindeki yatay çizgi hiçbir tercihe karşılık gelmez. 12 tavukta beyin 1 sn'de 1.5, 13 tavukta 4.5 sn'de beyin uyarıldı . (McCabe ve diğerleri, 1979.)
çıkrık ve onları tercih testinin yapılacağı ortama alıştırmak için onlara sabit bir sarı uyarıcı gösterin. İntraserebral stimülasyon toplam 5 saat sürmüştür, bu süre sonunda eş zamanlı seçim test aparatında tercih testi yapılmıştır. Aparatın her iki yanına görsel uyarıcılar (Şekil 9.1) yerleştirilmiştir . Her biri, biri 4,5 Hz'de, diğeri 1,5 Hz'de titreyen kırmızı bir ışıktı.
iki taraflı implante edilen elektrotlara sahip tavuklara ilişkin veriler , Şekiller 1 ve 2'de gösterilmektedir. 9.2. 1.5 Hz frekansta uyarılan tavuklar , 1 sn'de 1-5 kez yanıp sönen ışığı tercih ederken; 4,5 Hz elektrik stimülasyonu alan tavuklar , aynı frekanstaki flaşları tercih ettiler. Bilateral ek hiperstriatum veya ektostriatuma elektrot implante edilmiş 16 tavukta, stimülasyon sıklığının belirli bir görsel stimulus tercihi üzerinde hiçbir etkisi bulunmadı.
Aksesuar hiperstriatum doğrudan SMVG'ye uzanırken, ektostriatum multisinaptik bir yolla SMVG'ye bağlanır (Bölüm 11.1.1). Ancak bu iki görsel alanın elektriksel uyarımı Civcivlerin tercihini etkilemezken, PMVP'nin doğrudan uyarımı belli bir etki yaratmıştır. 1) görsel sistemden SMVG'ye tek bir giriş sinyalinin, alınan bilgilerin sonraki davranış testlerinde kullanılması olasılığını oluşturmak için yeterli olmadığı ve 2) sonuç olarak en az bir sistemden daha fazla sinyal gerektiği varsayılabilir. bilgileri kaydetmek ve sonra kullanmak için. PMCH'nin uyarılmasının iki varsayılan sistemi uyarması veya her iki sinyalin üzerinde birleştiği PMCH'deki postsinaptik elemanları doğrudan etkilemesi mümkündür. Beyin stimülasyonu üzerine yapılan deneylerin sonuçlarına dayanan yukarıda yapılan varsayımlar, Bölüm 1'de ileri sürülenlere yakındır. 6.4 (ayrıca bkz. şekil 6.6.4), ancak her durumda deneysel yaklaşım oldukça farklıydı. Elbette , PMVP'ye giriş sinyallerinden birini sağlayan ve damgalayıcı uyaranın pekiştirici veya motive edici etkisini uygulayan duyusal olmayan bir sistemin varlığını öne sürmek çekicidir, ancak görsel bir nesnenin etkileneceğine inanmak için herhangi bir neden var mı? böyle bir sistemi heyecanlandırmak?
Görsel bir uyaran, optik sinirde aktiviteye neden olarak talamus ve optik tavandaki nöronları ateşler. İkincisi, talamustaki bir sinaptik anahtar ile yolları ektostriatama gönderir (bölüm A.3.1). Görsel çatıdan, yollar beyin sapı da dahil olmak üzere beynin diğer bölümlerine götürür (Bradley ve diğerleri, 1985; bir inceleme için bkz. Pearson, 1972). Gövdenin orta kısmında yer alan hücre grupları sinir sisteminin birçok bölgesine yansıtılır. Memelilerde ve muhtemelen kuşlarda, ön beyne geniş uzantıları olan hücre gruplarından biri, hücreleri norepinefrin içeren mavi noktadır (Bölüm A.4). Görsel bir uyaran sadece önbeynin görsel bölgelerinde aktiviteyi indüklemekle kalmaz, aynı zamanda büyük olasılıkla beyin sapı hücrelerini de aktive eder (bkz. örneğin Watabe ve diğerleri, 1982). Bu nedenle, bir ön beyin nöronu görsel projeksiyon bölgesinden bir sinyal alırsa, aynı anda veya hafif bir gecikmeyle beyin sapından da bir sinyal alabilir. Böyle bir dolaylı etki, görsel bir uyaranın damgalama sırasında pekiştirici etkisini belirleyebilir mi?
Birkaç yıl önce Young ( 1963) , belirli öğrenme türlerinin veya hafıza kaydının pekiştireçlerden gelen sinyallere bağlı olduğunu öne sürdü . Griffith ( 1966), belleğin sinaps etkinliğindeki değişikliklere bağlı olduğu varsayımından yola çıkarak, takviyenin bu değişikliği destekleyebileceği bir hücresel mekanizma modeli önermiştir . Bununla birlikte, pekiştirme eyleminde hangi nöral yapı yer alır? Kety ( 1967), monoaminlerin memeli beynindeki geniş dağılımına dikkat çekti . Bu aminlerin seviyesini değiştiren maddelerin, özellikle katekolaminler olan dopamin ve norepinefrinin, insan ruh hali ve çeşitli hayvan davranışları üzerinde güçlü bir etkiye sahip olduğuna dikkat çekti . Bu veriler ışığında katekolaminlerin ve özellikle norepinefrinin affektif durumlarda ve beyin aktivasyon düzeyinin düzenlenmesinde önemli rol oynadığını ileri sürmüştür. Hem Kety (Kety, 1970, 1972) hem de Crow (Crow, 1968) , pekiştirmenin hafızanın temeli olarak kabul edilen sinaptik mekanizmalar üzerindeki etkisinin, hücreleri bulunan noradrenerjik sistemin nöronları tarafından gerçekleştirildiğini öne sürdü. beyin sapının orta kısmı. Kety ( 1970), "... beyindeki noradrenalinin etkisini tüketen veya bloke eden maddelerin konsolidasyonu yavaşlatması ve hafızayı bozması gerektiğini" tahmin etti . Serebral korteksi innerve eden noradrenerjik nöronların öğrenme mekanizmasının önemli bir bileşeni olduğu hipotezini test etmek için Anlezark ve arkadaşları ( 1973) farelerde locus coeruleus'a zarar verdi. Bu yaralanmalar, korteksteki norepinefrin konsantrasyonunun kontrol seviyesinin yaklaşık üçte birine düşmesine neden oldu . Bu tür yaralanmalara sahip farelerin, gıda takviyesi ile yaklaşma basit görevini öğrenme yeteneği ciddi şekilde bozulmuştur. Bununla birlikte, bu ümit verici başlangıca rağmen, en son araştırmalar , beyin norepinefrin seviyelerinde keskin bir düşüşün ardından öğrenme yeteneğinde genel bir bozulma olduğunu doğrulamamıştır (inceleme için bkz . Mason, 1984).
Beyin noradrenalin düşürücü müdahalelerin öğrenme üzerindeki etkisini yorumlamak zordur. Öğrenme üzerinde böyle bir etkinin tespit edildiği durumlarda, bu etki, uyanıklık, dikkat, konsantrasyon, motivasyon ve lokomotor aktivite seviyesindeki değişikliklere bağlı olarak davranış üzerindeki dolaylı etkilerden kaynaklanıyor olabilir - değişikliklerin yorumlanmasını zorlaştıran faktörler. hayvanın fizyolojik durumunu değiştiren hemen hemen her müdahaleden sonraki davranış (Bölüm 5). Norepinefrin seviyesindeki azalma ile davranıştaki değişikliklere de aracılık edilebilir. Örneğin, locus coeruleus'taki noradrenerjik nöronlar , adrenokortikotropik hormon ACTH'nin salınmasında yer alabilir (Amaral ve Sinnamon, 1977). Bu nöron sisteminin hasar görmesi , öncelikle strese neden olan koşullara uyum sağlama yeteneğini etkileyerek öğrenme yeteneğini etkileyebilir. Son olarak, nörotoksinler noradrenerjik sistemin fonksiyonlarını bozmak için kullanıldığında, davranışları etkileyen yan etkilere neden olabilirler. Beyindeki norepinefrin seviyesinde bir azalma ile belirli davranış türlerinin bozulmuş üretimine ilişkin verilerin çeşitli şekillerde yorumlanabileceği açıktır .
Benzer bir belirsizlik , beyindeki norepinefrin düzeyi ile yeni davranış biçimlerinin geliştirilmesi veya ezberlenmesi arasında bir ilişki ortaya koymayan deneylerin açıklamasında da mevcuttur . Örneğin, norepinefrin seviyesini düşürmeyi amaçlayan deneysel müdahaleler, norepinefrini ön beyinden asla tamamen ortadan kaldırmaz. Beyindeki plastik değişim seviyesi, görevin zorluğuna bağlı olarak değişiyorsa, o zaman beyindeki artık norepinefrin konsantrasyonu, nispeten basit görevlerde öğrenmeyi ve akılda tutmayı sürdürmek için yeterli, ancak daha karmaşık görevlerde yetersiz olabilir. Ek olarak, norepinefrin azalmasının sağlanma şekli, üretildiği yaş ve norepinefrin eliminasyonu ile öğrenmenin başlangıcı arasındaki sürenin uzunluğu, sonucu etkileyen önemli faktörler olabilir. Bear ve diğerleri ( 1983), norepinefrindeki kronik bir düşüşün etkilerinin daha sonra örneğin adrenerjik p -reseptörlerin yoğunluğundaki bir artışla büyük ölçüde maskelenebileceğine dikkat çekti (Sporn ve diğerleri, 1977; Harik ve diğerleri. , 1981). Bu nedenle, eğitim uygun zamanda gerçekleşirse, bu reseptörlerin yoğunluğundaki bir artış, norepinefrin sentezi seviyesindeki bir düşüşü telafi edebilir.
Öğrenme yeteneği ile beyindeki norepinefrin konsantrasyonu arasındaki ilişki hakkındaki veriler çelişkili olsa da, norepinefrinin nöronal plastisitedeki rolüne dair kanıtlar oldukça inandırıcıdır (Kasamatsu, 1983). Normal bir kedinin görsel korteksindeki nöronların yaklaşık %80'i her iki gözde de uygun uyarıya yanıt verir ve bu açıdan binokülerdir (Hubel ve Wiesel, 1962). Ancak, yaşamın erken bir döneminde bir gözün kapağı kapatılırsa, o zaman yavru kedilerin görme korteksindeki nöronlar yalnızca açık kalan göz tarafından kontrol edilmeye başlar ve çoğu ancak bu göz uyarıldığında uyarılır. (Wiesel ve Hubel/ 1963; Hubel ve Wiesel, 1970). Bu fizyolojik değişikliklere paralel olarak morfolojik değişiklikler de meydana gelir . Tek bir açık gözden sinyal ileten aksonlar, her iki gözü kullanan normal kedi yavrularına göre korteksin daha geniş alanlarını innerve eder (Wiesel, 1982 ). Göz hakimiyetindeki bu kayma, noradrenerjik ve dopaminerjik nöronları bozan bir maddenin, 6-hidroksidopamin (6-ODA) doğrudan görsel kortekse enjekte edilmesiyle önlenebilir (Kasamatsu ve diğerleri, 1979; Daw ve diğerleri, 1983; Bear ve diğerleri. ., 1983) . 6-ODA ile tedavi edilen yavru kedilerde kortikal plastisite , görsel kortekse doğrudan norepinefrin enjeksiyonu ile restore edilebilir (Kasamatsu ve diğerleri, 1979). Kasamatsu ve arkadaşlarının (Kasamatsu ve diğerleri, 1979) yaşlı kedi yavrularını veya yetişkin kedileri kullanarak yaptıkları çalışmalarda daha da önemli sonuçlar elde edilmiştir. Aynı zamanda göz kapağı dikilmesinin göz hakimiyetini etkileyebileceği sürenin yaşamın ilk üç ayı ile sınırlı olduğu dikkate alınmıştır (Hubel ve Wiesel, 1970): Bu süreden sonra tek taraflı göz kapağı dikilmesinin çok az etkisi vardır veya hiç yoktur. göz hakimiyeti. Kasamatsu , yaşlı hayvanlarda göz kapaklarını dikerek ve görsel kortekse norepinefrin enjekte ederek göz hakimiyetinde bir kayma elde edilebileceğini gösterdi. Başka bir deyişle, norepinefrin verilmesi yetişkin korteksinin plastisitesini eski haline getirmez .
1970) norepinefrinin sinapsları değiştirerek öğrenmeyi etkilediği yönündeki ilk önermeleri ile birlikte norepinefrinin oküler baskınlıktaki kaymayı etkileyen nöral mekanizmalarda rol oynadığını gösteren veriler, bizi bir azalmanın etkisini araştırmaya sevk etti. damgalama için beyindeki norepinefrin seviyesi ( Davies ve diğerleri, 1985). Böyle bir çalışma, yalnızca damgalamanın bazı yönlerinin öğrenme süreçlerini içermesi nedeniyle değil, aynı zamanda damgalamanın PMCG'deki sinaps morfolojisindeki değişikliklerle ilişkili olması nedeniyle umut vericidir (Bölüm 10.2). Önceden, katekolaminlerin tavuk beyninin belirli bölümlerinde bulunup bulunmadığını ve eğer öyleyse konsantrasyonlarının yaş ve görsel deneyimle değişip değişmediğini bulmamız gerekiyordu (Davies ve diğerleri, 1983).
Civciv beyninin farklı bölgelerindeki katekolamin seviyelerine yaş ve görsel deneyimin etkisi
Noradrenalin ve dopamin konsantrasyonları, tavuk ön beyninin üç bölümünde belirlendi: 1) Wulst; 2) esas olarak PMVG'yi içeren ön beynin medial kısmı; 3) esas olarak yükselen paleostriatumdan (paleostriatum boostatum) oluşan ön beynin bazal kısmı (bkz. Şekil P.Z). Farklı gruplardaki civcivlerden yumurtadan çıktıktan hemen sonra ve ayrıca 12, 22, 26, 29 ve 50 saat sonra doku parçaları alındı.Bir grup sürekli karanlıkta tutuldu; bu grubun sekiz tavuğu listelenen terimlerin her biri için kontrol olarak alındı. Diğer grup ise 21 saate kadar karanlıkta tutulduktan sonra 30 dakika tepeden ışıkla aydınlatıldı (s. 73), sonra tekrar karanlıkta bırakılarak 8 kişiden beyin dokusu örneği alındı. onların içindeyken

yumurtadan çıkma
Resim: 9.3. Karanlıkta tutulan tavukların ön beyinlerinin üç bölümünden alınan doku örneklerinde norepinefrin konsantrasyonları . Üstteki iki çizimin verileri , belirtilen parametreler için doğrusal regresyon denklemleriyle yeterince açıklanır. Bazal ön beyin için parametreler zamanla önemli ölçüde değişmez. Her puan, sekiz civciv grubunun ortalamasıdır. (Davies ve diğerleri, 1983.)
22 saat yaşlı Bu gruptaki diğer kuşlara 2 saat boyunca dönen bir kutu verildi. 8 tanesinin eğitimden hemen sonra, yaklaşık 26 saatlikken beyinleri alındı. Bu grupta kalan 24 civcivden, 12 tanesi eğitimden 3 saat sonra (29 saatlikken) ve 12 tanesi 24 saat sonra ( 50 saatlikken) katledildi.
Dopamin ve noradrenalin konsantrasyonlarında önemli hemisferik farklılıklar bulunmadı. Bazal ön beyinde maksimum ve Wulst'ta minimum olan dopamin konsantrasyonu yaş veya görme deneyimi ile değişmedi . Yumurtadan çıktıktan hemen sonra , ön beynin bazal kısmı norad-

Resim: 9.4. Görsel deneyimin civciv ön beynindeki norepinefrin konsantrasyonu üzerindeki etkisi . Diyagram, yumurtadan çıktıktan 22, 26, 29 ve 50 saat sonra alınan üç beyin bölgesinin (Wulst, medial ön beyin ve bazal ön beyin) birleşik skorunu temsil eder . Ortalama norepinefrin seviyesi, görme deneyimi olan (tek yumurtadan çıkan) tavuklarda, karanlıkta tutulan (çift yumurtadan çıkan) tavuklardan önemli ölçüde daha yüksekti (F' > 49 =5.57; P<0.05). Dikey parantez, iki araç arasındaki farkın standart hatasını gösterir. Her grupta 32 tavuk vardı. (Davies ve diğerleri, 1983.)
en yüksek konsantrasyonda renalin (1 g ağırlık başına 53.13 ± 7.85 ng ) ve ön beynin medial kısmı - en düşük seviyede (26.09 ± 3.75 ng/g). Yaşla birlikte, Wulst ve medial ön beyinde norepinefrin konsantrasyonu arttı (Şekil 9.3), ancak bazal beyindeki norepinefrin seviyeleri önemli ölçüde değişmedi.
İstatistiksel analiz, tüm Bölgeler bir araya toplandığında, görsel deneyim alan civcivlerde norepinefrin konsantrasyonunun, karanlıkta tutulan civcivlere göre önemli ölçüde daha yüksek olduğunu gösterdi (Şekil 9.4). Bu fark her üç bölüm ve farklı yaş grupları için yaklaşık olarak aynıydı. Sonuç olarak, yapay bir nesnenin damgalamaya neden olan etkisi , civciv telensefalonunda norepinefrin konsantrasyonunda genel bir artışa neden oldu. Daha önce, civcivlerin iki saat boyunca dönen bir kutuyla tanıştıktan sonra onu yeni bir nesneye tercih ettikleri gösterilmişti (örneğin bkz. Bölüm 5.2). Bu nedenle kutuyu gören tavukların kutuya bir bağ oluşturduğunu varsayabiliriz.
Orijinal çalışmanın sonuçları bazı umutları doğurdu, ancak bu umutlar büyük değildi. Dopamin konsantrasyonunun yaştan veya görsel deneyimden bağımsız olması , başlangıç seviyesi yeterli olabileceğinden, damgalama sürecine katılma olasılığını dışlamadı . Yaş ve görme deneyimi ile norepinefrin konsantrasyonundaki değişiklikler de bu aminin herhangi bir özel şekilde damgalamada yer aldığı anlamına gelmez. Ancak bu olumlu sonuç norepinefrine odaklanmamızı sağladı. Bu tür tavuklarda damgalamanın gelişip gelişmeyeceğini öğrenmek için beyindeki seviyesini bir şekilde azaltmak gerekiyordu .
Noradrenerjik nöronları etkileyen bir nörotoksin olan N-(2-kloroetil)-M-etil-2-bromobenzilamin hidroklorürün (DSP-4) beyindeki adrenalin bezlerinin bazı işlevlerinin araştırılmasında yararlı olduğu gösterilmiştir (Ross ve ark . ., 1973; Ross, 1976) . DSP-4'ün sıçanlara uygulanmasından sonra , kortekste ve merkezi sinir sisteminin diğer kısımlarında norepinefrin konsantrasyonunda uzun süreli bir düşüş meydana gelir (Ross, 1976; Jaim-Etcheverry ve Zieher, 1980). Jonsson ve diğerleri ( 1981) , fare beynindeki norepinefrin seviyeleri üzerinde DSP-4'ün benzer bir etkisini tanımlamıştır. Bu araştırmacılar, DSP-4'ün farelerde ve sıçanlarda noradrenerjik terminallerin nispeten seçici yıkımına neden olduğu sonucuna vardılar . Bu ve diğer sebeplerden dolayı (s. 205), bu maddeyi azalmış beyin norepinefrin düzeylerinin damgalama üzerindeki etkilerini incelemek için kullandık ( Davies ve diğerleri, 1985).
norepinefrin ve damgalama
Yumurtadan çıktıktan hemen sonra civcivler iki gruba ayrıldı. Deney grubuna (48 tavuk) damıtılmış suda bir DSP-4 çözeltisi enjekte edildi. Kontrol grubuna (ayrıca 48 tavuk) sadece distile su enjekte edildi. Enjeksiyonlardan sonra tüm civcivler karanlık inkübatöre alındı ve 30 saat sonra ikinci bir enjeksiyon yapıldı. 60 saat sonra, civcivler 30 dakika boyunca aydınlatıldı, sonra bir tekerleğe yerleştirildi ve 1 saat boyunca dönen bir kırmızı kutu veya doldurulmuş tavuk ile sunuldu. Tercih, ardışık bir testte 2 saat sonra ölçülmüştür . Yaklaşık 1 saat sonra civcivlerin sallanan boncuğa yaklaşma ve gagalama tepkilerini göstermeleri için geçen süre kaydedildi (bkz. Şekil 5.7, E).
Test sonunda her gruptan 12'şer adet civciv sakrifiye edildi ve hızlı bir şekilde beyinleri çıkarıldı. Bölümde listelenen üç alan. 9.2, her iki hemisferden disseke edildi. Daha önce herhangi bir departmanda konsantrasyonda asimetri bulunmadığından,
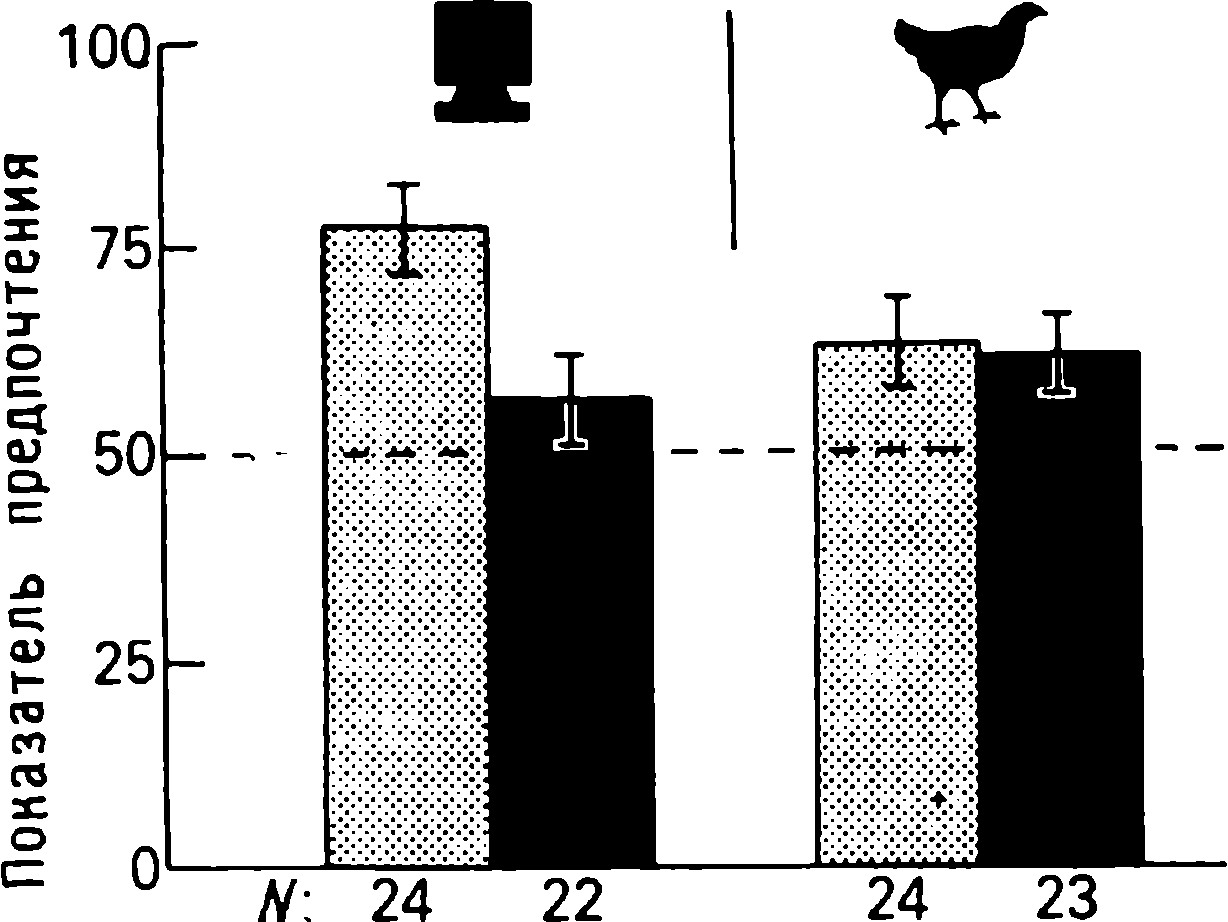
Resim: 9.5. DSP-4'ün tercih endeksi üzerindeki etkisi. Gösterilenler, kontrol grubu (noktalarla kaplı çubuklar) ve DSP-4'ün (siyah çubuklar) tanıtılmasından sonra tavuklar için ortalama değerlerdir. Kırmızı kutu ile eğitilen kontrol civcivlerinin tercih puanı 50'den önemli ölçüde büyüktü (P<0.001). DSP-4'ün tanıtılması, doldurulmuş tavukla eğitilmiş tavuklarda tercih puanını önemli ölçüde etkiledi ve bu tavuklarda ve kontrol grubundaki tercih puanları, 50'den önemli ölçüde yüksekti (P<0.02). (Davies ve diğerleri, 1985.)
norepinefrin ile dokuları birleştirildi ve biyokimyasal analizleri yapıldı.
Kırmızı kutuya maruz bırakılan kontrol civcivleri , bu uyaran için belirgin bir tercih geliştirdiler (Şekil 9.5). Bununla birlikte, DSP-4 uygulanan civcivlerin kutu tercih puanı , kontrol civcivlerinden önemli ölçüde düşüktü ve rastgele seviyeden önemli ölçüde farklı değildi. Buna karşılık, doldurulmuş tavuk göstererek eğitilen tavuklarda, DSP-4 grubunun puanı, kontrol grubunun puanı ile hemen hemen aynıydı. Tercih testi sırasında, DSP-4 ile tedavi edilen civcivler kontrol civcivleri kadar aktifti ve sallanan bilyenin gagalama gecikmesinde onlardan farklı değildi (bkz. Şekil 5.7e).
DSP-4 , incelenen üç ön beyin bölgesindeki (Şekil 9.6) adrenalin yuvalarının konsantrasyonunda yaklaşık %65 oranında önemli bir azalmaya neden oldu . Norepinefrin konsantrasyonundaki azalma , farklı nesneler kullanılarak eğitilen tavuklarda aynıydı .
Tercih puanları ile norepinefrin konsantrasyonu arasında ilişki olup olmadığını belirlemek için her bölüm için veriler ayrı ayrı incelenmiştir. Kırmızı kutu maruziyeti altında eğitilen tavukların orta (P<0.05) ve bazal (P<0.01) ön beyinlerinde anlamlı bir pozitif korelasyon bulundu, ancak Wulst'ta (P<0.2) bulunmadı. 14-140 eğitimli civcivler için tüm korelasyonlar
200
Kumaş Örneği: W & B
Resim: 9.6. DSP-4'ün ön beyindeki norepinefrin konsantrasyonu üzerindeki etkisi. Wulst (W), medial (M) ve bazal (B) ön beyinden alınan doku örnekleri için ortalama değerler verilmiştir . Çift taramalı sütunlar kontrol gruplarıdır; tek taranmış kolonlar, DSP-4 ile tedavi edilen tavuklardır . Bu maruz kalma, kontrol ile karşılaştırıldığında ön beyindeki norepinefrin konsantrasyonunda önemli bir azalmaya yol açtı (tüm bölümler için P <0.01). DSP-4 etkisinin eğitim sırasında kullanılan uyarana önemli bir bağımlılığı yoktu (F IıI7 = 0.26). (Davies ve diğerleri, 1985.)
doldurulmuş bir tavuğu gösterirken güvenilmezdi (tüm bölümler için P >>0.2).
Bu deneyin sonuçları, DSP-4'ün eğitimde kullanılan nesnenin doğasına bağlı olarak civcivin davranışı üzerinde farklılaştırılmış bir etkiye sahip olduğunu göstermektedir. Ancak bu sonuçlar başka bir şekilde açıklanabilir. DSP -4'ün doldurulmuş tavuk tercihini değiştirememesi "minimum seviye" etkisinden kaynaklanıyor olabilir. Bu varsayım doğrultusunda elde edilen sonuçlar aşağıdaki gibi açıklanabilir. DSP-4 yalnızca normalde yüksek ortalama tercih puanlarına sahip olan civcivleri etkiler. Bu tür göstergeler, kontrol tavuklarında kırmızı bir kutu ile sunulduklarında bulundu. Bu performans düzeyi, doldurulmuş bir tavukla sunulduğunda kontrol tavukları tarafından elde edilememiştir ; kutu eğitimli kontrol grubuna göre anlamlı derecede düşüktü (P<0.05). Bu nedenle, DSP-4, kırmızı kutu ile eğitilen civcivlerde tercihi etkiledi, ancak kukla ile değil.
bağlı olarak "minimum seviye" nin etkisini ve DSP-4'ün farklılaştırılmış eylemini ayırmak için ikinci bir deney gerçekleştirildi. Bu hariç, önceki seride olduğu gibi aynı şekilde planlandı
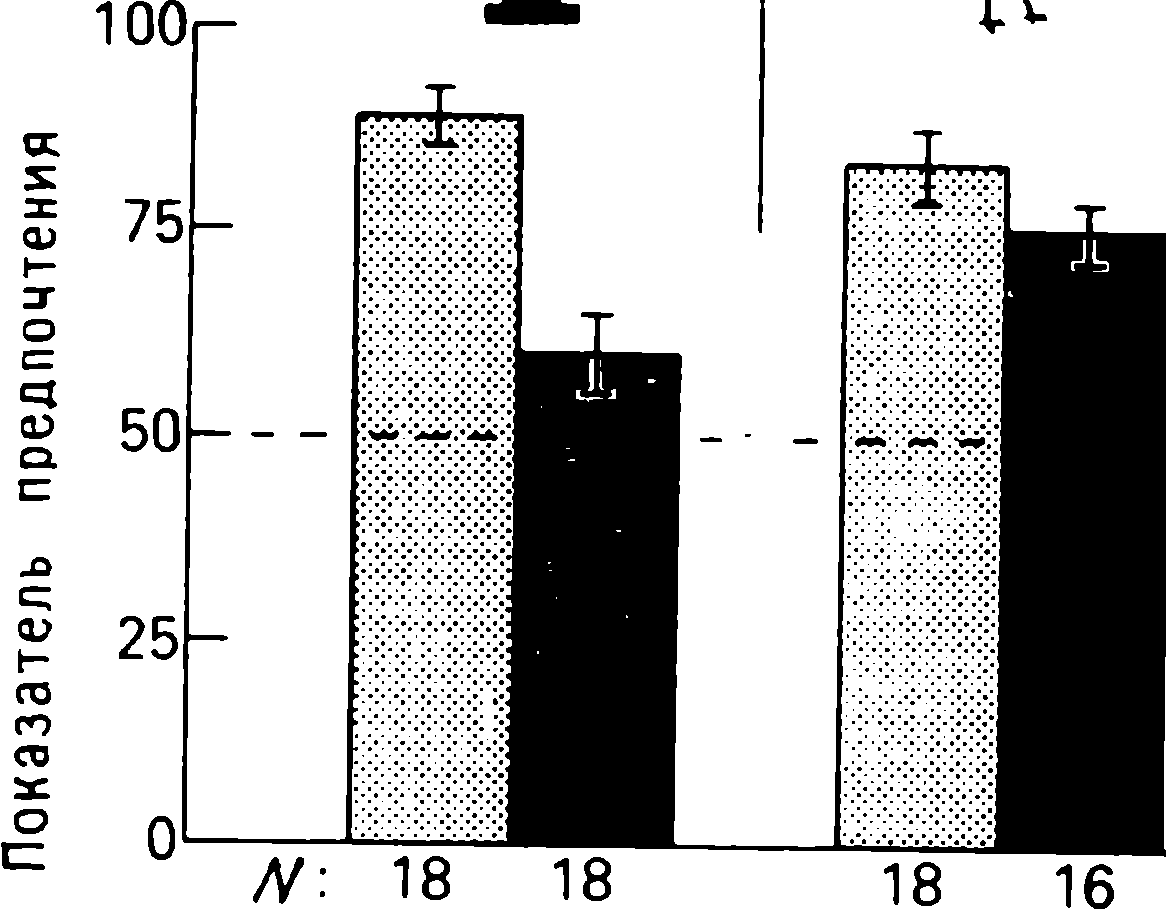
Resim: 9.7. DSP-4'ün tercih endeksi üzerindeki etkisi. Gösterilenler, kontrol grubu (noktalarla kaplı çubuklar) ve DSP-4 tedavi grubu (siyah çubuklar) için ortalama değerlerdir. İki kontrol grubu arasında anlamlı bir fark yoktu ve her iki puan da anlamlı olarak 50'nin üzerindeydi (P < 0.001). DSP-4 enjekte edilen ve doldurulmuş tavuk kullanılarak eğitilen grup için ortalama puan , kontrolden önemli ölçüde farklı değildi ve 50'den önemli ölçüde yüksekti (P < 0.001). Kırmızı kutu kullanılarak eğitilen DSP-4 ile tedavi edilen grup için ortalama puan , karşılık gelen kontrol grubundan (P<0.001) ve teşhirde eğitilen DSP-4 ile tedavi edilen gruptan önemli ölçüde daha düşüktü . <0,02); 50'den önemli ölçüde farklı değildi. (Davies ve diğerleri, 1985.)
eğitim sürecinde her iki nesnenin de gösterilmesi sırasında annenin arama kaydına yer verilmiş olması. Bu prosedür, görsel bir uyarana damgalamayı hızlandırır. Ayrıca antrenman süresi 80 dakikaya çıkarıldı. Bu modifikasyonların etkili olduğu kanıtlanmıştır (Şekil 9.7). Kırmızı kutu ve korkuluk gösterisi eğitimi alan kontrol gruplarının ortalama tercih puanları aynı bulunmuştur. Her iki gösterge de rastgele seviyeden önemli ölçüde farklıydı. Aynı şey, DSP-4 enjekte edilen ve onlara doldurulmuş tavuk sunularak eğitilen tavuklar için de geçerliydi; bu grubun göstergesi kontrol değerlerine yakındı. Buna karşılık, kutu gösterisinde eğitilen DSP - 4 grubu için medyan tercih puanı : 1) kontrol grubununkinden önemli ölçüde daha düşüktü; 2) doldurulmuş hayvana maruz kalma sırasında eğitilen DSP-4 grubununkinden önemli ölçüde daha düşüktü ; 3) rastgele seviyeden önemli ölçüde farklı değildi. Bu sonuçlar “minimum seviye” hipotezini çürütüyor çünkü iki kontrol grubunun yüksek ortalama puanları önemli ölçüde farklı değildi.
Özetleyelim. DSP-4 alan ve eski DSP-4 eğitimi alan grupta tercih puanları önemli ölçüde azaldı .
14* kırmızı kutunun konumu. Norepinefrin seviyesi, ön beynin tüm kısımlarında eşit olarak azaldı. Medial ve bazal ön beyindeki norepinefrin konsantrasyonu , yalnızca kırmızı kutu ile eğitilmiş tavuklarda tercih puanları ile pozitif korelasyon gösterdi .
incelenen yanıtı ikincil olarak etkileyen davranışın bazı gizli yönlerini etkilemenin sonucu mu olduğunu belirlemek önemlidir . Nitekim norepinefrin düzeylerini düşürmek için kullanılan bazı maddelerin davranış üzerinde çok net bir etkisi vardır. Kasamatsu ve Pettigrew'in (Kasamatsu ve Pettigrew, 1979) deneylerinde 6-ODA'nın beynin ventriküllerine girmesine, yalancı öfke nöbetleri, kasılmalar, kendini tutamama ve zorunlu dönüş eşlik etti. DSP-4 ile tedavi edilen tavuklarda bu tür patolojik reaksiyonlar gözlemlemedik. Bu tavuklar, tercih testlerinde kontrollerden daha az aktif değildi; DSP-4 ayrıca sallanan boncuğun gagalama gecikmesini de etkilemedi . Ancak DSP-4 , farklı nesnelerle eğitilen civcivlerin tercihlerinde aynı etkiyi gösterseydi, bu sonuca pek çok davranış değişikliğinden hangisinin neden olduğunu anlamak kolay olmayacaktı (s. 203-204). Aslında bu madde, yalnızca kullanılan iki uyarandan biri ile yetiştirilen tavukların tercihini güçlü bir şekilde etkilemiştir . Bu nedenle, herhangi bir spesifik olmayan davranışsal etki yoluyla damgalamayı etkilemesi olası değildir .
, esas olarak PMGF içeren medial ön beyindeki norepinefrin konsantrasyonu ve tercihler arasında pozitif bir korelasyon gözlendi . Bu sonuç, SMVP'nin yapay nesneler üzerine baskı yapmadaki rolünü bir kez daha doğrulamaktadır (bkz. Bölüm 8). Bu grup tavuklarda, tercih indeksleri ile bazal ön beyindeki noradrenalin konsantrasyonu arasında da pozitif bir korelasyon vardı . Bu doku alanı, esas olarak hareketin düzenlenmesinde rol oynayabilecek bir yapı olan yükseltilmiş bir paleostriatum içeriyordu ( Karten ve Dubbeldam, 1973; Rieke, 1980). Bununla birlikte, kırmızı kutu kullanılarak eğitilen tavuklarda bu alanda bulunan değişikliklerin lokomotor aktivite ile ilgili olması olası değildir. Doldurulmuş tavuğu gören tavuklar daha az aktif değildi, ancak norepinefrin seviyesi ile bir korelasyon göstermediler. Bu korelasyonun damgalama süreciyle bir ilgisi olup olmadığı henüz net değil. Yükseltilmiş paleostriatum ile PMWG arasında karşılıklı bağlantılar vardır ( s. 273), bu yapıların bazı işlevsel etkileşimlerini mümkün kılar (Bölüm 7.3). Wulst tercihi ile norepinefrin içeriği arasındaki korelasyon eksikliği, beyin hasarı deneylerinin sonuçlarıyla tutarlıdır: Kapsamlı Wulst yıkımı, civcivlerin damgalama sırasında görsel nesneler için bir tercih oluşturma ve sürdürme yeteneğini bozmaz (Bölüm 5).
Yukarıda tartışılan çalışmanın sonuçları (Davies ve diğerleri, 1985), norepinefrinin tavukların yapay bir nesnenin özelliklerini öğrenme sürecine katılımını bir dereceye kadar doğrulamaktadır . Ancak, bu onay sınırlıdır. Her şeyden önce, norepinefrinin kortikal plastisitedeki özel rolü sorgulanır (Sillito, 1983; Bear ve Singer, 1986). Shaw ve Sinader ( 1984), monoküler olarak yoksun kedi yavrularında göz hakimiyetindeki kaymaların , görsel korteksin L -glutamat ile perfüzyonu ile bloke edilebileceğini gösterdi. Bu etkinin noradrenerjik sistem üzerindeki bir etki yoluyla mı aracılık ettiği veya bu maddelerin her ikisinin de kortikal plastisiteyi bağımsız olarak etkileyip etkilemediği bilinmemektedir . Sinapstaki plastik değişikliklerin, yalnızca pre- ve postsinaptik nöronların deşarjları çakıştığında meydana gelmesi mümkündür (Hebb, 1949). Hem glutamat (Shaw ve Cynader , 1984) hem de norepinefrin (Kasamatsu ve Heggelund, 1982) kortikal nöronların aktivitesini değiştirir . Böylece bu maddeler kortikal nöronlardaki deşarj dizisini bozarak plastik değişiklikleri önleyebilir . Bu, norepinefrinin nöronal plastisitede fizyolojik bir rol oynama olasılığını dışlamaz . Örneğin , normal hayvanlarda norepinefrin, plastik değişiklikler için gerekli koşullara katkıda bulunarak nöronal aktiviteyi stabilize edebilir (ayrıca bkz. Bölüm 10.3.4 ve s. 249). DSP-4'ün doğasından şüphe etmek için başka nedenler de var . Bu maddenin damgalamayı norepinefrin düzeyini etkileyerek değil, norepinefrin konsantrasyonundaki değişikliklere paralel fizyolojik değişikliklerle etkilemesi olasıdır. DSP-4'ün, L-glutamat gibi , PMCG'deki ve tavuk beyninin diğer bölümlerindeki nöronların aktivitesini doğrudan bozması da mümkündür . Bu olasılıkları çürütecek hiçbir veri yoktur. Öğrenme sırasında sunulan uyaranın doğasına bağlı olarak DSP-4'ün damgalama üzerindeki farklı etkisi , PMCG hasarının damgalama üzerindeki etkisine benzer (Hinde ve McCabe, 1984). Bu yapının tahrip olması , kırmızı kutu ile sunulan tavuklarda damgalamayı keskin bir şekilde bozdu, ancak doldurulmuş tavuk ile sunulan tavuklarda çok daha zayıf bir etki yaptı. Civcivlerin bir kutu ve doldurulmuş bir tavukla eğitilmesinin sonuçlarının farklı mekanizmalara sahip olabileceğini düşündüren diğer kanıtlar , aşağıdaki deney serilerinden gelmektedir (Bolhuis ve diğerleri, 1986).
Testosteron ve damgalama
Testosteron ile tedavi edilen genç erkeklerin dikkati daha az dağılır ve testosteron içermeyen civcivlere göre çeşitli uyaranlara daha agresif tepki verir (Andrew, 1972; Archer , 1974). Bir dizi deneyi özetleyen Andrew ve diğerleri (Andrew ve diğerleri, 1981), testosteronun hafıza ve dikkat oluşumunda ortak bir mekanizma üzerinde hareket edebileceğini öne sürdüler. Dikkat ve hafıza damgalama için gereklidir. Bu hormon damgalamayı etkiler mi? Birkaç yazar (James, 1962; Balthazart ve de Rycker, 1979) testosteronun civcivlerde yaklaşma tepkisini etkilediğini göstermiştir, ancak bu araştırmacılar, hormonun damgalamanın doğrudan bir göstergesi olan tercih üzerindeki etkisini analiz etmemiştir . Bu nedenle, testosteronun damgalama üzerindeki etkisini araştırdık (Bolhuis ve diğerleri, 1986).
Bu deneylerde civcivler karanlıkta yumurtadan çıktı; kısa bir süre sonra cinsiyetleri belirlendi (Davies ve Raupe, 1982). Her tavuğa, fıstık ezmesi (deney grubu) veya tek başına fıstık ezmesi (kontrol grubu) içinde çözülmüş tek bir deri altı testosteron enjeksiyonu yapıldı . Uygulanan testosteron dozları 1, 3 veya 5 mg idi. Testosteron enanthate, bu ilacın uzun süreli etkisi olduğu için kullanıldı; kan seviyesi en az 24 saat sabit kalır ( Andrew, 1983). Her testosteron dozu eşit sayıda kadın ve erkeğe uygulandı. Enjeksiyondan sonra civcivler her zaman olduğu gibi kodlandı, böylece her civcivin geçmişi sonraki eğitim, test ve hormon tayini yapanlar tarafından bilinmiyordu .
Enjeksiyondan yaklaşık 16 saat sonra civcivler 22-27 saatlikken 30 dakika ışıklandırıldı, ardından dönen tekerleklere yerleştirildi ve 2 saat dönen bir kutu veya doldurulmuş hayvanla sunuldu. Eğitim sonunda civcivler tekrar 30 dakika karanlık inkübatöre alındı. 2 saat sonra sıralı bir testteki tercihleri ölçüldü ve tercih puanları daha önce açıklandığı gibi hesaplandı (s. 104). Daha sonra tekrar karanlık bir inkübatöre yerleştirildiler, kanları alındı ve plazmaları ayrıldı. Plazma testosteron konsantrasyonu, radyoimmunoassay ile ölçüldü (Yalow, 1980). Testosteron, plazmada bulunan tek erkek cinsiyet hormonu (androjen) değildir; kullanılan yöntem 5a-dihidrotestosterona duyarlıydı (%22.4 çapraz reaksiyon). Kısaca, diğer yazarların yaptığı gibi (örneğin, Tanabe ve ark., 1979), belirleme sonuçlarına plazma "testosteron" konsantrasyonu olarak atıfta bulunacağız.
Plazma testosteron konsantrasyonlarında veya tercih puanlarında cinsiyet farklılıkları yoktu. Kontrol grubundaki plazma testosteron seviyelerinde cinsiyet farklılıklarının bulunmaması, Tanabe ve arkadaşlarının sonuçlarıyla tutarlıdır (Tanabe ve diğerleri, 1979). Bu araştırmacılar, kadınlarda ve erkeklerde testosteron konsantrasyonlarının yaşamın ilk 4 haftasında farklılık göstermediğini gösterdi.
Eksojen testosteronun ortalama tercih puanları üzerindeki etkisi, kullanılan uyaranın doğasına bağlıydı. Bu nedenle doldurulmuş tavuk ve kırmızı kutu ile sunulan tavuklara ait veriler ayrı ayrı işlenmiştir. Önce doldurulmuş bir tavuğun gösteriminden elde edilen sonuçları ele alalım . Tüm bu grup için genel ortalama puanlar 61.74±2.11 idi. Tercih indeksi , eksojen testosteronun etkisi altında değişir: deney grubunda 64.52 ± 2.32 ve kontrol grubunda 54.76 ± 4.30; göstergelerdeki farklılıklar istatistiksel olarak anlamlıdır (P<0.05). Kontrol grubu için puanlar rastgele 50 seviyesinden önemli ölçüde farklı değildi. Tavuklara 2 saat boyunca doldurulmuş tavuk verildiği düşünülürse bu biraz garipti . Tercih puanları testosteron konsantrasyonuna göre sıralandığında kısmi bir açıklama sağlandı. Ortalama konsantrasyon 1.07 ng/ml idi. Testosteron konsantrasyonunun bu seviyenin üzerinde olduğu kontrol tavukları için ortalama tercihler 50'yi önemli ölçüde aştı (P<0.05) ve geri kalanı için 50'den önemli ölçüde farklı değildi.
Birlikte ele alındığında, bu sonuçlar doldurulmuş tavuklarla eğitilmiş civcivlerde tercih ile plazma testosteron konsantrasyonları arasında bir ilişki olduğunu göstermektedir. Bu, belirtilen iki gösterge arasında pozitif bir korelasyonun varlığı ile doğrulanmaktadır . Korelasyon, kontrol (Şekil 9.8, /4) ve deneysel (Şekil 9.8, Z>) tavuklarda ayrı ayrı ve bu gruplar birleştirildiğinde (Şekil 9.9, B) önemliydi. Bu nedenle, tercih puanları ve testosteron seviyeleri, fizyolojik testosteron konsantrasyonlarının hem içinde hem de dışında ilişkilidir. Tercih göstergelerinin kortikosteron düzeyine bağımlılığını gösteren regresyon çizgisinin eğimi (Şekil 9.8), deney grubu için önemliydi.

Plazma testosteron konsantrasyonu, ng/ml
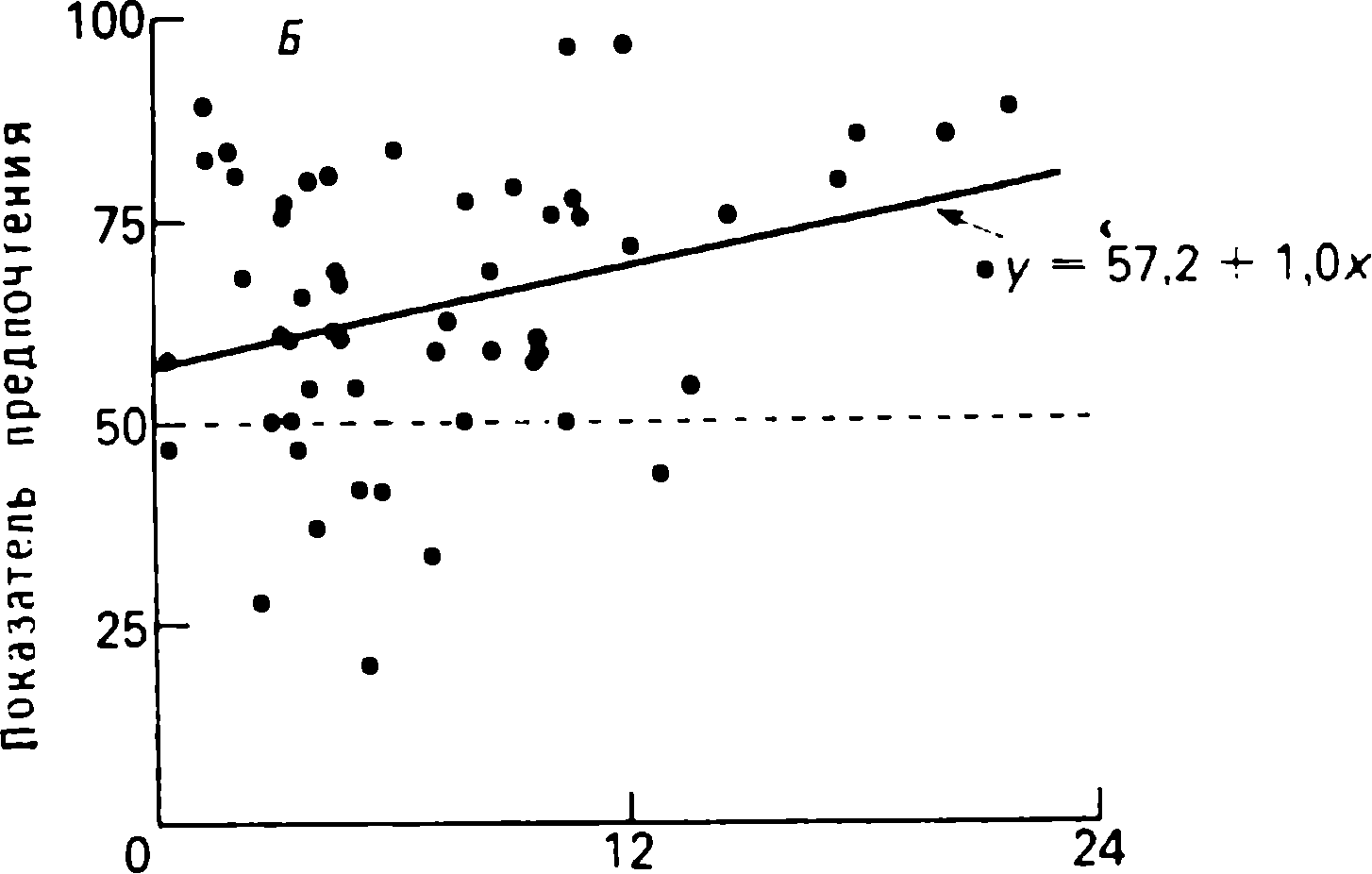
Plazma testosteron konsantrasyonu, ng/ml
Resim: 9.8. Doldurulmuş tavukla eğitilen civcivlerde tercih puanları ile plazma kortikosteron konsantrasyonu arasındaki ilişki. Verilere dayanarak, belirtilen parametrelerle doğrusal regresyonlar oluşturuldu. A. Kontrol grubu. Korelasyon katsayısı (rα=0.62; 20 serbestlik derecesi) Р<0.002'de anlamlıdır. B. Deney grubu. Korelasyon katsayısı (r «=0.30; 55 serbestlik derecesi) Р<0.02'de anlamlıdır. (Bolhuis ve diğerleri, 1986.)
kontrolden önemli ölçüde daha az (P<0.05). Bu nedenle , bu değişkenlerin bağımlılığını yüksek testosteron konsantrasyonu seviyelerinde eşitleme eğilimi vardır.
Tüm kırmızı kutu eğitimli civcivler için genel ortalama tercih puanı 70.14 ± 2.02 idi. Deney ve kontrol gruplarının göstergeleri arasında anlamlı bir fark yoktu. Bu kuşlarda, ister ayrı ayrı ister birlikte alınsın , testosteron konsantrasyonu ile tercih puanları arasında anlamlı bir ilişki yoktu .
tüm tavuklar için ortalama tercih indeksi (70.14±2.02), doldurulmuş tavuk ile sunulan tüm tavuklardan (61.74±2 ,11) anlamlı derecede yüksekti (P<0.01). Tercih puanları 100'den yüksek olamayacağı için kutuyu gören civcivlerin korelasyonunun olduğu ancak bu grup için tercih puanlarının teorik maksimuma, yani “tavana” yakın olması nedeniyle bunun maskelendiği söylenebilir . etkisi görüldü.. Bu olasılığı araştırmak için, bu grup için tercih puanları sıralandı, en yüksek olanlar atıldı ve geri kalanlar için ortalama değer hesaplandı. Bu prosedür, ortalama değer, eğitim sırasında doldurulmuş hayvanla sunulan tavukların değerlerine yaklaşana kadar tekrarlandı ve bu, 23 tavuğun verileri elenerek elde edildi. Bu kısıtlı grup için ortalama tercih puanı 61.68±1.93 idi. Bununla birlikte, bu durumda da anlamlı bir korelasyon bulunmadı (Şekil 9.9,/4). Regresyon çizgisinin eğimi, doldurulmuş hayvan kullanılarak eğitilen tüm tavuklardan önemli ölçüde daha düşüktü ve sonuç olarak, uyarandaki farklılıklara bağlı olarak veriler arasındaki tutarsızlık, ortalama tercihler eşitlendikten sonra bile devam etti.
Eksojen testosteronun civciv davranışı üzerindeki etkisi spesifik değil mi, örneğin sensorimotor koordinasyonu veya aktivasyon seviyelerini etkiliyor mu, yoksa tercih oluşumunda daha doğrudan mı yer alıyor? Testosteronun bir tercih testi sırasında aktivasyon seviyesini veya el-göz koordinasyonunu etkilemesi olası değildir , çünkü tanıdık ve yeni nesne yaklaşma tepkileri testosteron seviyeleri ile önemli ölçüde ilişkili değildir. Bununla birlikte, hormonun spesifik olmayan etkisine karşı en güçlü argüman, seviyesinin sadece eğitim sırasında doldurulmuş tavukla sunulan tavuk gruplarında tercihlerle ilişkili olmasıdır.
incelenen iki değişken arasında nedensel bir ilişkinin kanıtı değildir . Ayrıca, nedensel bir ilişki olsa bile , korelasyon nedenin ne olduğunu ve sonucun ne olduğunu göstermez. Doldurulmuş tavuk için güçlü bir tercih testosteron salgılanmasına yol açar mı, yoksa doldurulmuş tavuk için güçlü bir tercih geliştirmek için yüksek testosteron seviyeleri gerekli midir? Emzik tercihi testosteron sekresyonunu arttırıyorsa, manken kontrol civcivlerindeki α konsantrasyonu kutulu gruplara göre daha yüksek olmalıdır.
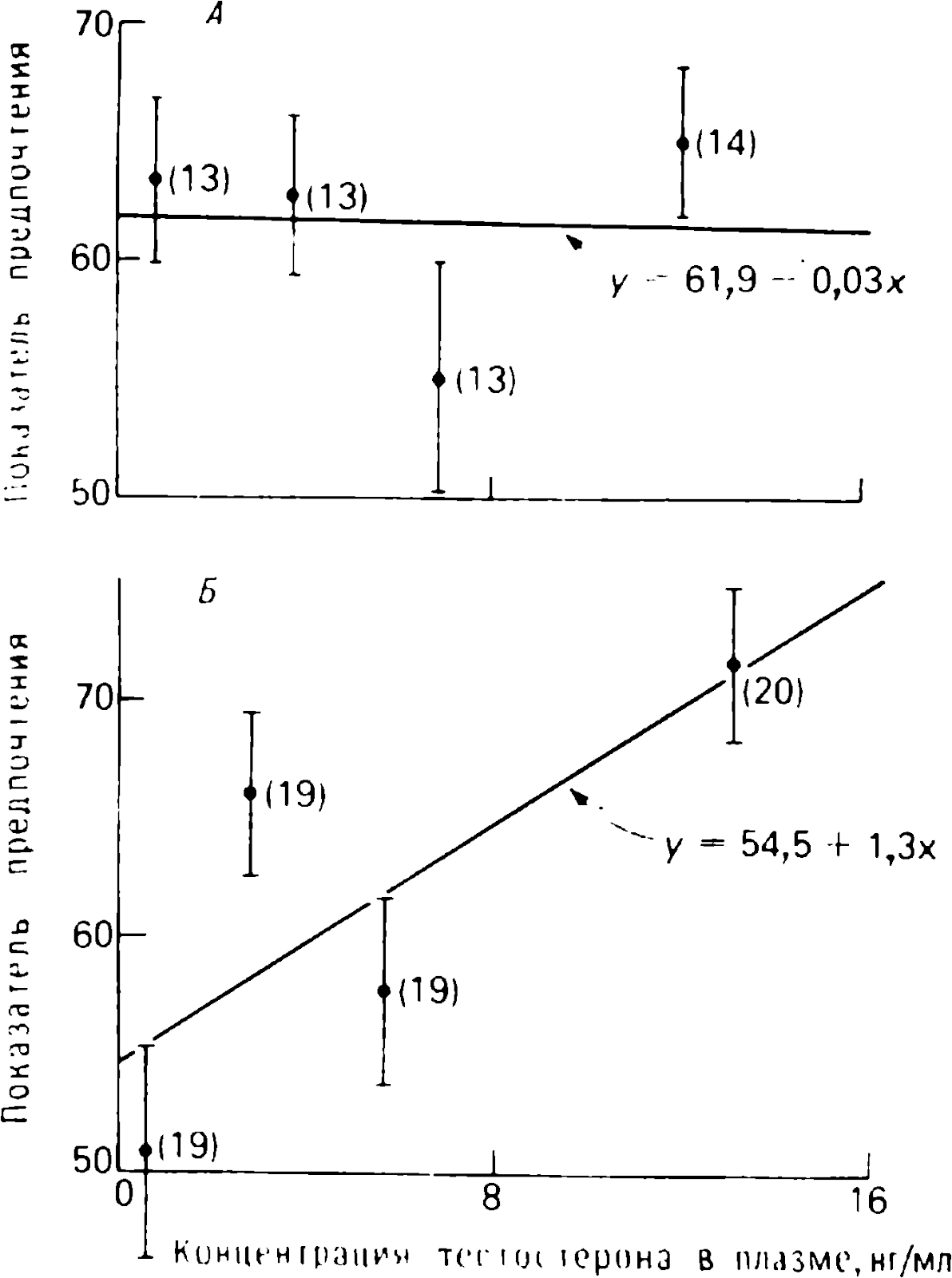
Resim: 9.9. Tercih puanı ile plazma testosteron konsantrasyonu arasındaki ilişki . Kontrol ve deney gruplarına ait veriler birleştirilir. Kutu eğitimli civcivlerin ortalaması (metne bakın) doldurulmuş tavuk grubununkiyle karşılaştırıldı. Testosteron konsantrasyonlarının dağılımı çeyreklere ayrıldı ve ortaya çıkan dört alt grup için ortalama değerler grafikler üzerinde çizildi. Tavuk sayısı parantez içinde verilmiştir. Dikey çizgiler : ± 1 SE _ _ serbestlik, P <<0.01). A ve B'de gösterilen çizgilerin eğimleri önemli ölçüde farklıdır (P<0.05). (Bolhuis ve diğerleri, 1986.)
Bununla birlikte, iki tavuk grubundaki ortalama plazma konsantrasyonu önemli ölçüde farklılık göstermedi. Tercih puanlarının testosteron konsantrasyonuna bağımlılığını açıklamanın doğrudan bir yolu , ekzojen testosteron uygulamasıdır. Aynı zamanda plazmadaki testosteron seviyesi önemli ölçüde artar ve aynı zamanda doldurulmuş tavuk tercihleri artar . Böylece, bu tavuk grubu için, tercih ölçümlerini etkileyen testosteron konsantrasyonu ile nedensel bir ilişkiyi gösteren veriler elde edildi .
modülasyonu yoluyla belirli davranışsal tepki türlerinin oluşumunda testosteronun etkisi hakkındaki varsayım tamamen yeni değildir. Örneğin, kanaryada şarkı söylemeyi ve diğer seslendirmeleri kontrol eden beynin belirli bölgeleri tanımlanmıştır (Nottebohm ve diğerleri, 1976; Nottebohm, 1980; ayrıca bkz. s. 161). Zebra ispinozlarında (Peophila gutta ta) benzer yapılar şarkı söylemeyi kontrol eder (Gurney ve Konishi, 1980). Şarkı, erkek ispinozlarda kastrasyonla basitleştirildi ve ekzojen androjenlerin eklenmesiyle eski haline getirildi (Pröve, 1974; Arnold, 1975). Bu kuşta şarkı söylemekten sorumlu bazı yapılar , stosteron veya metabolitlerini biriktirir (Arnold, 1980). Bu çalışmalar , androjen hedef hücrelerinin kanaryalar ve ispinozlarda şarkı öğrenmede yer aldığını ve bu işlev için androjenlerin gerekli olduğunu göstermektedir (Arnold, 1981; Nottebohm, 1981).
çift ayrışma
Sadece eğitim sırasında kırmızı bir kutuya maruz kalan tavuklarda DSP-4, tercih tepkilerini güçlü bir şekilde etkiledi. Böylece, eğitimde kullanılan nesneye bağlı olarak DSP-4 etkilerinin basit bir ayrışması vardı . Testosteron çalışmaları, etkilerin basit bir ayrışmasına başka bir örnek sağlar. Sadece eğitim sırasında doldurulmuş tavuk sunulan tavuklarda testosteron tercih puanlarını etkiledi. "Çifte ayrışma" nın bir örneği olan bu iki etkinin birleşimi (Teuber, 1955; Shallice, 1979; Jones, 1983), bu iki tercih biçiminin altında yatan sinirsel mekanizmaların en azından bazılarının farklı bir fizyolojik yapıya sahip olduğunu göstermektedir. doğa.
Yorumlar ve akıl yürütme
Bölümde açıklanan deneyler. 9.1 ve 9.3, duyusal yollardaki dürtü aktivitesinin, civcivin bu aktiviteye neden olan yapay bir nesneye bağlanması için yeterli olmadığını göstermektedir. Diğer faktörlerin de dahil edilmesi gerekir . Eğitim sırasında kırmızı bir kutuya maruz kalan civcivler için sınırlayıcı faktör, ön beyindeki norepinefrin konsantrasyonu olabilir. Belirli sınırlar içinde , DMAH bölgesindeki bu aminin konsantrasyonu ne kadar yüksekse, kırmızı kutu tercihi o kadar güçlüdür. Bu sonuçlar, Kety (1970, 1972) ve Crow'un (Crow, 1968; Crow ve Arbuthnott, 1972) uyaranın güçlendirici etkisine noradrenerjik sistemin aracılık ettiği fikirleriyle tutarlıdır . Bu sonuçlar ayrıca, damgalama sırasında bir nesne hakkındaki bilginin yalnızca depolama sisteminin hem duyusal hem de motivasyonel sistemlerden sinyaller alması durumunda sabitlendiği hipoteziyle tutarlıdır (Bölüm 6.4).
Norepinefrinin damgalamadaki olası rolünü gösteren veriler elbette diğer maddelerin de katılımını dışlamaz (s. 214). Ayrıca , norepinefrin seviyesinde keskin bir düşüş koşulları altında bazı sinir yapılarının modifikasyon olasılığını da dışlamazlar . Aslında, doldurulmuş tavuk tercihinin geliştirilmesinde rol oynayan tam da böyle bir sistemdir . Bu nesneyle sunulan tavuklar , beyin norepinefrin seviyelerinin %65 oranında azalmasına rağmen kutuyu tercih ettiler . Bununla birlikte, bu gruptaki tercih, plazma testosteron konsantrasyonu ile ilişkilidir ve muhtemelen bu faktör tarafından sınırlandırılmıştır.
Doldurulmuş tavuğun tercihi, tavuklar onu hiç görmemişken bile gelişir. Bunu yapmak için , yaklaşık 20 saatlik yaştaki tavukları, ışıkta veya karanlıkta bir veya iki saat dönen bir tekerleğe yerleştirmek yeterlidir. 24 saat sonra test edildiklerinde yine de doldurulmuş tavuğu kırmızı kutuya tercih edecekler (Bölüm 8.2). Ancak 2 saat sonra bu tercih henüz kendini göstermiyor. Bu sonuçlar için olası bir açıklama, bu prosedürün , doldurulmuş tavuk için tercih sağlayan bazı özel sistemleri işlevsel hale getirmesidir. Bu tercih plazma testosteron seviyeleri ile sınırlı olabileceğinden, plazma fonksiyonuna bu hormonun aracılık etmesi mümkündür. Bu varsayımın bir takım sonuçları vardır. Dönen bir çarkta biraz zaman geçiren civcivler , karanlık bir kuluçka makinesinde kalan civcivlere göre çarkın üzerindeyken daha fazla manipülasyon, belki daha fazla hareket ve farklı bir duyusal deneyim yaşarlar . Bu deneyimler birlikte veya ayrı ayrı plazma testosteron düzeylerini etkiler mi? Şu anda, bu konuda doğrudan verilerimiz yok. Bununla birlikte, Krulik ve diğerleri (Kruhlich ve diğerleri, 1974) , laboratuar farelerinde ön hipofiz bezindeki bir dizi hormonun salgılanmasının, hayvanların bir odadan diğerine taşınması dahil olmak üzere çeşitli prosedürlerin etkisi altında değiştiğini göstermiştir. . Bunlardan biri - luteal düşürücü hormon - testosteron salgılanmasını uyarır. Luteinizan hormonun salgılanması bu transferden sonra yaklaşık bir saat kadar yükselir ve kontrol seviyelerine geri dönmez. Artışı, testosteron konsantrasyonunda bir artışa yol açabilir. Civcivler dönen bir tekerleğe yerleştirildiğinde benzer olaylar yaşanıyorsa , testisler veya yumurtalıklar tarafından testosteron salgılanmasında ortaya çıkan artış (Tanabe ve diğerleri, 1979) , doldurulmuş tavuk tercihini sürdüren varsayımsal sistemi tetiklemek için yeterli olabilir. bu işlem birkaç saat sürecek olsa da.
ortaya çıkan tercihe bu tür bir mekanizmanın yanıt vermesi mümkündür (Bölüm 8.2). Benzer bir mekanizma, civcivler bir çıkrığa yerleştirildiğinde ve karmaşık bir desenle veya doldurulmuş hayvanın kendisiyle sunulduğunda tercih gelişiminin hızlanması gibi diğer sonuçları açıklayabilir mi? Böyle bir sistemde duyusal sinyaller ve testosteron seviyelerinin etkileşim içinde olması mümkündür . Belirli bir testosteron seviyesinin arka planına karşı, nöronlar arasındaki işlevsel bağlantı, optimal duyusal uyarımla , yetersiz olandan daha güçlü olabilir. Karmaşık görsel uyaranlar , dağınık aydınlatmadan daha verimli hücre aktivasyonuna neden olabilir, ancak doldurulmuş bir tavuk daha da etkili olmalıdır. Nöronların etkileşimi ne kadar güçlüyse, bir tercih testinde doldurulmuş tavuk gösterildiğinde nöronlar o kadar aktif bir şekilde boşalırlar . Buna göre, yaklaşma reaksiyonunu kontrol eden motor nöronlara efferent sinyallerin akışı daha güçlü olacak ve bu reaksiyonun kendisi daha aktif olacaktır.
işlevsel olarak açılması nasıl gerçekleşebilir? Örneğin, önerilen sistemin nöronlarının başlangıçta yalnızca belirli bir testosteron seviyesinin arka planına karşı işlevsel bağlantılar kurması mümkündür (Bölüm 10.3.7). Nöronlar daha sonra , doldurulmuş bir tavuğun doğasında bulunan uyaran kombinasyonları da dahil olmak üzere, belirli uyaran kombinasyonlarına (örneğin, bir kafa ve bir gözün görünümünün bir kombinasyonu; bkz. Hinde, 1954; Candland, 1969; Coss, 1972) seçici olarak yanıt verebilir. Bu sistemin nöronları, yaklaşma tepkilerini kontrol eden motor yapılara bağlıysa, o zaman doldurulmuş bir hayvanın işlevsel içerme sürecinden sonra sunulması, tavuk bu nesneyi daha önce hiç görmemiş olsa bile, seçici bir yaklaşma tepkisine neden olacaktır.
Beyin hasarı ile ilgili deneylerin sonuçları, PMVP'nin böyle özel bir sistemin parçası olmadığını göstermektedir (Bölüm 8.1 ve 8.4.1). Bununla birlikte, PMVG'nin dışında bulunan varsayılan tanıma sisteminin işlevsel olarak açılmasından sonra, nasıl açılırsa çalıştırılsın , tavukların türlerinin belirli bir temsilcisine yaklaşması ve özelliklerini ezberlemesi mümkündür. PMVP'nin bu sürece dahil olduğu varsayımını zaten ifade ettik (s. 179) ve ilgili verileri bunun lehine gösterdik (Bölüm 8.4.2). PMVG'nin rolü, örneğin bireysel özelliklerin kaydedilmesi gibi, özel bir yaklaşım sistemi için belirli uyaranların ayrıntılarını sabitlemekten ibaret olabilir. Bu bakış açısına göre, işlevsel aktivasyonu testosteronun katılımıyla gerçekleşen özel bir sistem, tavuğu aynı türün temsilcilerine yönlendirir. Bundan sonra, nöronlar arasındaki fonksiyonel bağlantılar. optimal uyaran tarafından uyarıldıkça ve anneyi tanımak için gereken bilgi PMCH depolama sistemine girildiğinden sistemler daha verimli hale gelebilir .
PMVP'de eser oluşumu için noradrenalin gerekliyse , bu süreç ön beyindeki norepinefrin seviyesindeki bir azalma ile bozulmalıdır. Bu amaçla DSP-4'ün kullanıldığı iki deneyde , eğitim sırasında tavuk kafası verilen kuşların ortalama tercih puanları biraz azaldı. Bu deneylerin birinde (Şekil 9.7) bulunan %8.52±4.73'lük azalma istatistiksel olarak anlamlıydı (/=1.80, P<0.04). Bu azalma, PMVP'nin yok edilmesiyle gözlemlenene yakındır: yıkım eğitimden sonra yapılırsa, doldurulmuş tavuğun tercih edilmesi , kontrol grubuna kıyasla yaklaşık %11 oranında azalır (Tablo 8.1).
Elbette, söz konusu iki maddenin , testosteron ve norepinefrin, karşılık gelen nesneler gösterilen tavuklarda yatkınlık ve tercihin tezahürünü sınırlayan tek faktör olduğu varsayılamaz. Tıpkı "genel motivasyon durumu" kavramının yerini davranışı etkileyen çok sayıda etkileşimli faktöre bırakması gibi (Hinde, 1970, 1982), çok çeşitli fizyolojik faktörlerin çeşitli davranış biçimlerine kısıtlamalar getirmesi beklenebilir. örneğin, beslenme, civciv ve daha yaşlı kuşlarda - yırtıcılardan kaçınmak ve cinsel bir eş seçmek. Ve hafıza da dahil olmak üzere herhangi bir işlevin uygulanmasında birden fazla faktör söz konusu olabilir (s. 249).
Özet
Duyusal yollardaki aktivitenin, bilgilerin belleğe kaydedilmesi için yetersiz olduğu verileri dikkate alınır. Dahili durumla ilişkili faktörler , izlerin oluşumuna kısıtlamalar getirir . Noradrenalin ve testosteronun damgalama sürecindeki rolü analiz edildi. Eğitim sırasında kırmızı bir kutu ile sunulan tavuklarda PMVG içeren ön beyin bölgesindeki norepinefrin konsantrasyonunun hafızayı sınırladığı ortaya çıktı. Doldurulmuş bir tavuğu göstermek için eğitilmiş civcivler için bu nesneye yönelik tercih, plazma testosteron konsantrasyonu ile ilişkilidir . Açıklanan deneylerin koşulları altında , testosteron kırmızı kutu tercihini etkilemedi. Bu iki nesnenin tercih edilmesinin altında yatan mekanizmaların en azından bazılarının farklı fizyolojik yapıya sahip olduğu sonucuna varılmıştır. Sec.'de açıklanan doldurulmuş tavuk için kendiliğinden oluşan tercihin olduğu varsayılmaktadır .
testosteronun etkisi altında aktive edilebilen belirli bir sistemin aktivitesinin bir sonucudur.
EDEBİYAT
Amaral DG, Sinnamon HM Locus coeruleus: Merkezi bir noradrenerjik çekirdeğin nörobiyolojisi, Progr. Neurobiol., 1977, 9, 147-196.
Andrew RJ Tanıma süreçleri ve davranışı, testosteronun kalıcılık üzerindeki etkilerine özel referansla, Adv. Çalışma Davranışı, 1972, 4, 175-208.
Andrew RJ Östradiol ve testosteronun genç evcil civcivde dikkat dağınıklığı ve hafıza oluşumu üzerindeki spesifik kısa gecikmeli etkileri. İçinde: Yüksek omurgalılarda hormonlar ve davranış (cds. J. Balthazart, E. Pröve ve R. Gilles), 1983, s. 463-473. Springer-Vergie^, A. Berlin.
Andrew RJ, Clifton PG, Gibbs ME Evcil civcivlerde testosteron ile öğrenmenin etkinliğinin arttırılması, J. comp. fizik. PsychoL, 1981, 95, 406-417.
Anlezark GM, Crow TJ, Greenway AP Bilateral locus coeruleus lezyonlarından sonra bozulmuş öğrenme ve azalmış kortikal norepinefrin, Science, 1973, 181, 682-684.
Archer J. Testosteronun civcivlerin ilgisiz ve ilgili yeni uyaranlarla dikkatlerinin dağılması üzerindeki etkileri, Anim. Behav., 1974, 22, 397-404.
Arnold AP Zebra ispinozlarında (Poephila guttato), J. exp. Zool., 1975, 191, 309-326.
Arnold AP Zebra ispinozu beynindeki hormon birikimindeki cinsiyet farklılıklarının nicel analizi: metodolojik ve teorik sorunlar, J. comp. Neurol., 1980, 189, 421-436.
Arnold AP Omurgalı davranışının kontrolünde steroid hormon etkisinin mantıksal seviyeleri, Am. Zool., 1981, 21, 233-242.
Balthazart J., de Rycker C. Baskılama durumundaki civcivlerin davranışları üzerinde androjenlerin ve östrojenlerin etkileri, Z. Tierpsychol., 1979, 49, 55-64.
Bateson PPG, Wainwright AAP Daha önce ışığa maruz kalmanın evcil civcivlerde baskı süreci üzerindeki etkileri, Behavior, 1972, 42, 279-290.
Ayı MF, Şarkıcı 1F. Görsel kortikal plastisitenin asetilkolin ve noradrenalin ile modülasyonu, Nature, Londra., 1986, 320, 172-176.
Bear MF, Paradiso MA, Schwartz M, Nelson SB, Carnes K.M. , Da niels JD Kitty'de iki katekolamin tükenme yöntemi, plastisite üzerinde farklı etkiler sağlar, Nature, Londra., 1983, 302, 245-247.
Bolhuis J. L, McCabe BJ, Horn G. Androjenler ve baskı. Evcil civciv Behav'da testosteronun evlatlık tercihleri üzerindeki farklı etkileri. Nörobilim., 1985.
Bradley P., Davies DC, Horn G. Evcil civcivde (Gallus domesticus) hyperstriatum ventrale'nin bağlantıları, J. Anat., 1985, 140, 577-589.
Candland DK Evcil tavuğun sosyal baskınlık düzenini sürdürmede kullandığı yüz bölgelerinin ayırt edilebilirliği, J. comp. fizik. PsychoL, 1969. 69, 281-285.
Coss RG Göz benzeri şemalar: davranış üzerindeki etkileri. Doktora tezi, Reading Üniversitesi, 1972.
Crow TJ Kortikal sinapslar ve takviye: bir hipotez, Nature, Londra., 1968, 219, 736-737.
Crow TJ, Arbuthnott G. W, Memeli Merkezi sinir sisteminde katekolamin içeren nöronların işlevi. Nature new Biol., 1972, 238, 245-246.
Davies DC, Payne JK Yumurtadan çıkma sırasında civciv cinsiyet oranlarındaki değişim. Animasyon Behav., 1982, 30, 931-932.
Davies D.C., Horn G., McCabe BJ Evcil civcivlerde telesefalik katekolamin düzeylerindeki değişiklikler. Yaş ve görsel deneyimin etkileri, Devel. Brain Res., 1983, 10, 251-255.
Davies DC, Horn G. t McCabe BJ Noradrenalin ve öğrenme: nörotoksin DSP4'ün evcil civciv Behav'da damgalama üzerindeki etkileri. Neurosci., 1985, 99, 652-660.
Daw NW, Rader RK, Robertson TW, Ariel M. 6-hidroksi dopaminin yavru kedi çizgili korteksinde görsel yoksunluk üzerindeki etkileri, J. Neurosci., 1983,3, 90-914.
Griffith JS Belleğin doğasına ilişkin bir teori, Nature, Londra., 1966, 211, 1160-1163.
Gurney ME, Konishi M. Zebra ispinozlarında beyin ve davranışın hormon kaynaklı cinsel farklılaşması, Science, 1980, 108, 1380-1383.
Harik SI, Duckrow RB, LaManna JC, Rosenthal M., Sharma VK, Banerjee SP Locus ceruleus lezyonunun neden olduğu kronik noradrenerjik denervasyon için serebral kompanzasyon: reseptör bağlanmasının, izoproterenol kaynaklı adenilat siklaz aktivitesinin ve oksidatif metabolizmanın geri kazanımı, J. Neurosci. , 1981, 1, 641-649.
Hebb DO Davranışın organizasyonu, John Wiley, New York, 1949.
(Fringilla coelebs) yıldırma davranışında gösterildiği gibi, kısmen doğuştan gelen bir tepkinin gücündeki değişiklikleri yöneten Hinde RA Faktörleri. I. Tepkinin niteliği ve seyrinin incelenmesi, Proc. R. Soc., 1954, B142, 306-331.
Hynd R. Hayvan davranışı. 1975, M., "Barış".
Hinde RA Etolojisi: doğası ve diğer Bilimlerle ilişkileri. Fontana, Londra 1982.
Hinde RA, McCabe BJ Önyargılar ve tercihler. Lezyonların civciv beynine damgalanması üzerindeki etkiler, Anim. Behav., 1984, 32, 288-292.
Hubel DH, Wiesel TN Alıcı alanlar, binoküler etkileşim ve kedinin görsel korteksindeki işlevsel mimari, J. Physiol. Londra., 1962, 160, 106-154.
Hubel DH, Wiesel TN Yavru kedilerde tek taraflı göz kapatmanın fizyolojik etkilerine duyarlılık dönemi, J. Physiol. Londra., 1970, 206, 419-436.
Jaim-Etcheverry G., Zieher LM DSP4: yetişkin ve gelişmekte olan farelerin noradrenerjik nöronları üzerinde nörotoksik etkileri olan yeni bir bileşik, Brain Res., 1980, 188, 513-523.
James H. görsel titreme ile damgalama: testosteron siklopentilpropiyonat, Anim'in etkileri. Behav., 1962, 10, 341-346.
Jones GV On double disociation of function, Neuropsychologia, 1983, 21, 397-400.
Jonsson G., Hallman H., Ponzio F., Ross S. DSP4 (N-(2-kloroetil)-N-etil-2-bromobenzilamin) - Merkezi ve periperhal noradrenalin nöronları için yararlı bir denervasyon aracı, Eur. J. Pharmac., 1981, 72, 173-188.
Karten HJ, Dubbeldam JL Güvercindeki (Columba livia) paleostriatal kompleksin organizasyonu ve izdüşümleri, J. comp. Neurol., 1973, 148, 61-90.
Kasamatsu T. Kedi görsel korteksinde Merkezi norepinefrin Sistemi tarafından korunan nöronal plastisite , Progr. Psychobiol., fizyol. Psychol., 1983, 10, 1-112.
Kasamatsu T., Heggelund P. Noradrenalinin iyontoforezi sırasında görsel uyarıma kedi görsel korteksinde tek hücreli yanıtlar, Exp. Brain Res., 1982, 45, 317-327.
Kasamatsu T., Pettigrew JD 6-hidroksidopamin, J. komp. Neurol., 1979, 185, 139-162.
Kasamatsu T., Pettigrew JD t Ary M. Norepinefrinin lokal mikroperfüzyonu ile görsel kortikal plastisitenin restorasyonu, J. comp. Neurol., 1979, 185, 163-182.
Kety SS Biyojenik aminlerin merkezi fizyolojik ve farmakolojik etkileri ve bunların davranışla ilişkileri. İçinde: Nörobilimler: Bir çalışma programı (editörler G.C. Quarton, T. Melnechuk ve F.O. Schmitt), 1967, s. 444-451. Rockefeller University Press, New York.
Kety SS Merkezi sinir sistemindeki biyojenik aminler: uyarılma, duygu ve öğrenmedeki olası rolleri. İçinde: Nörobilimler: ikinci çalışma programı (ed. F. O. Schmitt), 1970, s. 324-336. Rockefeller University Press, New York.
Kety SS Korteksin adrenerjik sistemlerinin öğrenmedeki olası rolü, Res. Yayınlar Ass. Res. sinir ment. Dis., 1972, 50, 376-386.
Kruhlich L., Hefco E., Oku C. B. Akut stresin normal sıçanda LH, FSH, prolaktin ve GH salgılanması üzerindeki etkileri, istatistiksel değerlendirmeleri üzerine yorumlar, Neuroendocrinology, 1974, 16, 293 -311.
McCabe BJ, Horn G., Bateson PPG Ritmik hiperstriatal stimülasyonun civcivlerin görsel titreme tercihleri üzerindeki etkileri, Physiol. Behav., 1979, 23, 137-140.
Mason ST Katekolaminler ve davranış, 1984. Cambridge University Press, Cambridge.
Nottebohm F. Kuşlarda ses öğrenimi için beyin yolları: ilk 10 yılın gözden geçirilmesi, Progr. Psychobiol., fizyol. Psychol., 1980, 9, 86-124.
Nottebohm, F. Tüm mevsimler için bir beyin: kanarya beynindeki şarkı kontrol çekirdeklerinde döngüsel anatomik değişiklikler, Science, 1981, 214, 1368-1370.
Nottebohm F., Stokes TM, Leonard CM Kanaryadaki şarkının merkezi kontrolü, Serinus canarius, J. comp. Neurol., 1976, 165, 457-486.
Pearson R. Kuş beyni, 1972, Academic Press, New York.
Pröve E. Der Einfluss von Kastration und Testosteronsubstitution auf das Sexual- verhalten mânnlicher Zebrafinken (Taeniopygia guttata castanotis Gould), J. Om. Lpz., 1974, 115, 338-347.
Okuyucu TA Kedinin görsel korteksindeki dopamin, noradrenalin ve serotoninin etkileri, Experientia, 1978, 34, 1586-1587.
Rieke GK Kainik asit lezyonları: hareket bozukluklarının incelenmesi için bir model, Physiol. Behav., 1980, 24, 683-687.
Ross SB N-2-kloroetil-N-etil-2-bromobenzilamin hidrokloridin sıçan beyni ve kalbindeki noradrenerjik nöronlar üzerindeki uzun süreli etkileri, Br. J. Pharmac., 1976, 58, 521-527.
Ross SB, Johansson JG, Lindborg B., Dahlbom R. Bisiklete binme tesisleri. I. Adrenerjik bloke edici etkiye sahip tersiyer N-(2-bromobenzil)-N-haloalkilaminler, Açta Pharm. suecica, 1973, 10, 29-42.
Shallice T. Nöropsikolojik araştırma ve hafıza sistemlerinin fraksiyonlanması. İçinde: Hafıza araştırmasına ilişkin perspektifler (ed. L.-G. Nilsson), 1979, s. 257-277.
Shaw C., Cynader M. Kortikal aktivitenin bozulması, monoküler olarak yoksun kedi yavrularında oküler baskınlık değişikliklerini önler, Nature, Londra., 1984, 308, 731-734.
SilUto AM Görsel kortekste Plastisite, Nature, Londra., 1983, 303, 477-478.
Sporn JR, Wolfe V. B., Harden T. K-, Molinoff PB Sıçan serebral korteksinde aşırı duyarlılık: 6-hidroksidopaminin norad renergic sinapslarında pre-' ve postsinaptik etkileri , Molec. Pharmac., 1977, 13, 1170-1180.
Tanabe Y. t Nakamura T., Fujioka K., Doi O. Embriyonik dönemde testisler, yumurtalıklar ve adrenal bezler tarafından seks steroid hormonlarının üretimi ve salgılanması
140 ve genç tavuk (Gallus domesticus). Gen. komp. Endocrinal., 1979, 39, 26-33.
Teuber H.-L. Fizyolojik psikoloji, A Rev. Psychol., 1955, 6, 267-296.
W at abe K, Nakai K, Kasamatsu T. Eat locus coeruleus'ta norepinefrin içeren nöronlara görsel afferentler, Exp. Brain Res., 1982, 48, 66-80.
Wiesel TN Doğum sonrası görsel korteksin gelişimi ve çevrenin etkisi, Nature, Londra., 1982, 299, 583-591.
Wiesel TN, Hubel DH Tek gözde Görüşten yoksun kedilerin çizgili korteksinde tek hücre tepkisi, J. Neurophysiol., 1963, 26, 1003-1017.
Yalow RS Radioimmunoassay, A. Rev. biyografiler. Bioengng., 1980, 9, 327-345.
Young JZ Nöral hafıza sistemlerinin bazı esasları. Eylem sonuçlarının sinyallerini düzenleyen ve adresleyen eşleştirilmiş merkezler, Nature, Londra., 1963> 198, 626-630.
ÖĞRENME SÜRECİNİN FİZYOLOJİK VE MORFOLOJİK SONUÇLARI
PMVP'nin özel rolünün ilk belirtileri, protein ve RNA sentezi öncüllerinin kullanıldığı deneylerde elde edildi. Bu deneylerin temeli , eğer bellek hücre büyümesiyle ilişkiliyse, o zaman muhtemelen protein ve RNA sentezinde de değişiklikler olması gerektiği varsayımıydı. Deneylerin sonuçları, damgalamanın öncüllerin makromoleküllere dahil edilmesindeki değişiklikleri ortaya çıkardığını gösterdi . Urasil birleşimindeki değişiklikler , öğrenmenin basit bir yan etkisi olarak açıklanamaz ; Bu değişikliklerin yapay nesnelerin neden olduğu damgalama süreciyle yakından ilişkili olduğu ortaya çıktı. Radyootografik çalışmalar, bu değişikliklerin PMVP'de meydana geldiğini göstermiştir. Bu alanın iki taraflı olarak tahrip edilmesi, tavukların bu tür nesneler için bir tercih oluşturma veya daha önce edinilmiş bir tercihin izlerini tutma yeteneğini bozdu . Aşağıdaki bölümlerde sunulan bu ve diğer sonuçlar, PMIG'lerin damgalama , tanıma ve hafıza süreçlerinde belirleyici bir rol oynadığını göstermektedir . Ayrıca, sol PMVG'nin kendisinin, yaklaşma tepkilerini uyandıran belirli nesnelerin nöral modellerinin bir deposu olabileceğine dair basit bir hipotez öne sürüldü. Kuşkusuz, LPMVG, bozulmamış tavuklardaki izlerin tek deposu olamaz, ancak depolardan biri olarak işlev gördüğü deneysel verilerle doğrulanır (Bölüm 7.1). Dikkatle ve bu sonucu kabul ederek , orijinal biyokimyasal çalışmaların dayandığı varsayıma dönülebilir: bilginin kaydedilmesi nöronlarda morfolojik değişiklikler gerektirir. Elbette bu varsayım yanlış çıkabilir. Özellikle PMVG'de bulunan RNA metabolizmasındaki değişiklikler, hücrelerdeki morfolojik değişikliklerle ilişkili olmayabilir. Bunlar, örneğin bazı nöropeptitleri kodlayan haberci RNA'nın sentezindeki bir değişiklik gibi, daha incelikli bazı işlevsel yönleri yansıtabilirler . Bununla birlikte, aynı soru kalır: PMCH'de imprinting ile ilişkili morfolojik ve fizyolojik değişiklikler meydana gelir mi? Önce fizyolojik değişikliklerle ilgili verileri dikkate almak daha uygundur.
on beş*
Öğrenmenin Fizyolojik Sonuçları
uyarı olmadan kendiliğinden aktif kalırlar (Brown ve Hoth , 1979). Damgalama ile ilgili PMVG'nin fizyolojik etkilerini analiz etmenin ilk adımı olarak, damgalama uyaranına maruz kaldıktan sonra nöronların aktivitesini kaydettik (Raupe ve Horn, 1982, 1984). Kontrol için ek bir hiperstriatumda kayıt yapıldı.
Yaklaşık 21 saatlik civcivler 30 dakika boyunca dağınık ışığa maruz bırakıldı. İki saat boyunca 36 tavuğa dönen kırmızı kutu şeklinde bir uyarıcı ve diğer 18 tavuğa doldurulmuş tavuk verildi. Daha sonra karanlık inkübatöre geri döndüler. Yaklaşık 46 saat yaşına geldiklerinde 1 saat boyunca tekrar aynı nesne gösterildi.Genelde eğitim sırasında her civciv 3 saat boyunca dönen bir çarkın içindeydi.Tekerleğin dönüş sayısı değerlendirme için bir gösterge görevi gördü. nesneye yaklaşma etkinliği . Eğitim bitiminden 1 saat sonra civcivlere anestezi uygulandı . Beynin sol yarıküresine iki elektrot yerleştirildi: biri aksesuar hiperstriatuma, diğeri PMSH'ye. Elektrotlar , 250 um'lik bir adımla dikey olarak hareket ettirildi. Her hareketin sonunda spontan hücre aktivitesi 1 dakika süreyle kaydedildi. Her penetrasyonda bu türden en az üç kayıt yapılmıştır. Her hatta kaydedilen deşarjların ortalama sıklığı hesaplandı. Kayıt sırasında deneyciler tavuğun ne tür etkilere maruz kaldığını bilmiyorlardı . Kayıt tamamlandıktan sonra, türetme yeri işaretlendi . Her elektrotun izi, lekeli beyin bölümlerinden alınan eskizlerden yeniden oluşturuldu.
Kırmızı kutu ile sunulan 36 civcivde, 3 saatlik eğitim için toplam yaklaşım değerleri ile PMVP'deki ortalama spontan aktivite sıklığı arasında anlamlı bir korelasyon bulundu. Korelasyon negatifti , yani civciv kutuya ne kadar çok yaklaşmaya çalışırsa, PMSH'deki nöronal deşarjların sıklığı o kadar düşüktü (Şekil 10.1, L).
Aynı tavuklarda ek hiperstriatum kaydı yapıldığında farklı bir sonuç elde edildi . Anlamlı bir korelasyon bulunmadı (Şekil 10.1.5). Bu durumda korelasyon katsayısı , PMSH için karşılık gelen katsayıdan önemli ölçüde farklıydı . Bu nedenle, nöron aktivitesi ile yakınlık göstergeleri arasındaki ilişkiler , beynin farklı bölümleri için farklıydı.
Ayrıca her bölgedeki ortalama deşarj sıklığı ile <at- göstergeleri arasındaki korelasyon katsayılarını hesapladık.
Genel yakınlık göstergeleri
Resim: 10.1. Tavuk eğitimi sırasında nöron deşarjlarının seviyesi ile kırmızı kutuya yaklaşma tepkisi arasındaki ilişki. 3 saatlik eğitim sırasında ortalama spontan aktivite sıklığının yaklaşma aktivitesine bağımlılığı gösterilmiştir. Regresyon çizgileri en küçük kareler yöntemi kullanılarak oluşturulmuştur . 36 tavuğun her birinde, kayıt mikroelektrotları aynı anda SMVG'ye ve ek hiperstriatuma yerleştirildi. A. PMVG. Korelasyon katsayısı anlamlıydı (r=-0.54; 34 serbestlik derecesi; P<0.01). B. Ek hiperstriatum. Anlamlı bir korelasyon yoktur (r=-0.02; 34 serbestlik derecesi). İki korelasyon katsayısı arasındaki farklar anlamlı değildir (P<0.05). (Raupe, Değil, 1984.)
eğitimin 1. saatinde yakınlık. Korelasyon katsayısı PMAH için anlamlı (r = -0.50; serbestlik derecesi sayısı 34'e eşit; P<0.01) ve ek hiperstriatum için anlamsız (r = -0.004; serbestlik derecesi sayısı 34) olarak kaldı.
Doldurulmuş bir tavuğun gösterilmesiyle yapılan deneylerde , PMIG'deki ortalama nöron aktivitesi sıklığı ile yaklaşık göstergeler arasında ne eğitimin 1. saatinde ne de bir bütün olarak tüm süre boyunca anlamlı bir ilişki bulunmadı (Şekil 10.2, L). Bununla birlikte, her iki korelasyon katsayısı (her iki durumda da r = -0.07) aynı negatif yöne sahipti ve kutu kullanılarak eğitilen 36 civciv grubunda karşılık gelen korelasyon katsayılarından önemli ölçüde farklı değildi. Bu nedenle, doldurulmuş bir tavuğu 18 tavuktan oluşan daha küçük bir gruba göstermenin zayıf etkisi kolayca göz ardı edilemez. Ek hiperstriatumun nöronlarının ortalama deşarj sıklığı , genel yaklaşım göstergesi (Şekil 10.2, B) ve eğitimin 1. saati göstergesi ile ilişkili değildi.
PMSH'deki dürtü aktivitesi ile uyaranın doğasına yakınlık göstergeleri arasındaki ilişkinin açık bağımlılığı, örneğin, grubun eğitilmiş olması gerçeğiyle açıklanabilir.

Genel göstergeler, yaklaşımlar
Resim: 10.2. Nöron deşarjlarının seviyesi ile doldurulmuş tavuğa yaklaşma tepkisi arasındaki ilişki. Sonuçları Şekil l'de gösterilen deneydeki gibi tüm koşullar. 10.1. PMVH (A) ve ek hiperstriatum (B) için korelasyonlar istatistiksel olarak anlamlı değildir . (Raupe, Değil, 1984.)
doldurulmuş hayvan kullanımıyla, öğrenme sürecinde daha az aktifti . Ancak bunu destekleyecek herhangi bir veri bulunmamaktadır. Eğitimin ilk saatindeki yaklaşım tepkilerinin ortalama göstergesi ve 3 saatlik toplam gösterge , farklı nesneler kullanılarak eğitilen tavuk gruplarında önemli ölçüde farklılık göstermedi.
Yakınlık puanları ile PMCH hücrelerindeki deşarj seviyeleri arasındaki korelasyon üzerindeki damgalayıcı uyaranın farklı etkisi, tavukları kırmızı bir kutuya veya doldurulmuş tavuğa maruz bırakmanın farklı etkilerinin başka bir örneğidir (bölüm 8 ve 9). PMCG nöronlarının elektrofizyolojik analizi , kırmızı kutu bir nesne olarak kullanıldığında, bu yapının damgalama sürecindeki rolünü doğrular .
Elektrofizyolojik sonuçların yorumlanması, PMIG'de bilgi işleme için impuls aktivitesinde tespit edilen kaymaların önemi hakkında daha fazla veri elde edene kadar spekülatif kalacaktır. Muhtemel yorumlardan biri, daha düşük bir deşarj seviyesinin, kaydedilen nöronlar üzerinde daha güçlü inhibitör etkilerin varlığını gösterdiğidir. Bu tür engelleyici etkiler seçiciyse, o zaman tanıdık bir nesnenin neden olduğu etkinliği etkilemeden SMVP'deki yeni nesneler tarafından uyarılan etkinliği bastırabilirler .
Öğrenmenin morfolojik sonuçları
Beyinde bilgi depolamanın sinir bağlantılarındaki değişikliklere dayandığına inanılmaktadır (Tanzi, 1893; Cajal, 1911; Konorski, 1948; Hebb, 1949). Bu nedenle öğrenmenin morfolojik sonuçları incelenirken sinapslara dikkat edilmiştir. LMWH'nin bilgi depolamadaki rolüne ilişkin ilk çalışma sırasında (Bradley ve diğerleri, 1979, 1981), bu alanın işlevlerinde interhemisferik asimetri hakkında hiçbir veri yoktu . Yapay bir nesne tarafından etkinleştirilen bazı sinirsel mekanizmaların , doldurulmuş bir tavuk tarafından etkinleştirilenlerden farklı olabileceği de bilinmiyordu . Neyse ki, bu çalışmalarda PMVG'lerin her iki hemisferinden doku örnekleri alındı ve damgalama için uyarıcı olarak yapay bir nesne kullanıldı.
21 saatte karanlıkta yetiştirilen civcivler, 30 dakika boyunca dağınık ışıkla aydınlatıldı ve ardından bir baskılayıcı uyaranla sunuldu. Radyootografik çalışmalarda da benzer bir uyaran kullanıldı; tek fark, yukarı doğru hareket eden yarığın sarı yerine kırmızı olmasıydı. Daha sonra her iki grubun civcivlerine 20 dakika süre ile uyaran verilmiş ve yaklaşma tepkileri ölçülmüştür. Bu aktiviteye göre civcivler uygun çiftlere ayrıldı ve aktif olmayanlar deneyden elendi (s. 89-90). Her çiftten bir civciv artık eğitilmedi ve karanlık inkübatöre ("yetersiz" grup) geri gönderildi ve diğeri, aralarında 40 dakikalık aralıklarla ("aşırı eğitimli") 40 dakikalık üç periyoda bölünerek 120 dakika daha eğitildi. Grup). Bundan sonra, karanlık inkübatöre geri döndüler , kodlandılar ve her zamanki gibi sonraki tüm prosedürler kör bir şekilde gerçekleştirildi. Eğitimin başlamasından 6,5 saat sonra civcivler yaklaşık 30 saatlik olduklarında anestezi uygulandı. Bu zamana kadar, "yetersiz eğitim almış" kuşlar , eğitimin bitiminden sonra yaklaşık 4,5 saat boyunca karanlık bir inkübatörde tutuldu ; "yeniden eğitilmiş" grup için bu süre yaklaşık 3 saatti Beynin histolojik yapısını korumak için tavuklara bir fiksatif perfüze edildi. Daha sonra ön beyinden PMWG'nin bir kısmını içeren yaklaşık 1 mm kalınlığında enine bir kesit alındı. 1 mm kalınlığında PMVG bloğu izole edildi ve ultra ince kesitler alındı. Bir bloktaki sinaps örneklerini ölçmek için her onuncu dilim bir elektron mikroskobu kullanılarak incelendi. Mikrofotoğraflar sistematik olarak her 4. günde bir alınmıştır. Ve PMVP'nin her bloğundan 25 mikrofotoğraf çekildi. 22200 kez son büyütmede basıldılar ve kantitatif analiz için kullanıldılar. Sinaps, sinaptik keseciklerin varlığı, postyaptik sıkıştırma ve pre- ve post-aptik zarları ayıran bir boşluk ile kimyasal olarak tanımlandı (Şekil 10.3). Daire yüzdesini tahmin etmek için

Resim: 10.3. SMVG'deki kimyasal sinapsların bazı özelliklerini gösteren elektron mikrografları . Presinaptik elemanlar veziküller içerir. İki ok arasındaki elektron yoğun bölge , sinapsın aktif bölgesidir. Bu alan, postsinaptik mühür, presinaptik zardan dar bir yarıkla ayrılır. A. Akson terminali (presinaptik tomurcuk, 1) ile dendrit (2) üzerindeki omurga arasındaki sinaps . B. "Yol boyunca" akson ("geçiş", 3) ile dendritin gövdesi (4) arasındaki sinaps . (Bradley ve diğerleri, 1981.)
Hücre gövdeleri, sinaptik tomurcuklar ve dendritik dikenlerin kapladığı mikrograf alanında (Gray, 1959), örnekleme yöntemi kullanıldı (Weibel ve Bolender, 1973; Mayhew, 1979). Dikenlerdeki sinaps sayısının dendritik gövdelerdeki sinaps sayısına oranı da hesaplandı . Postsinaptik zarın karakteristik bir özelliği , sitoplazmanın yanında elektron yoğun malzemenin bulunmasıdır ( Reters ve diğerleri, 1976); bu postsinaptik sıkıştırmanın uzunluğunu ölçtük. Yedi "az öğrenilmiş" ve yedi "fazla öğrenilmiş" kuştan toplam 3711 sinaps analiz edildi. Veriler her tavuğun sol ve sağ PMVG'si için ayrı ayrı elde edildi . Her iki tavuk grubundan alınan iki PMVG'nin fotomikrografları en az iki kez karıştırıldı. Üzerindeki tüm ölçümler , araştırmacının fotoğrafta sunulan beyin yarımküresini veya tavukların davranışsal deneyimlerini bilmeden gerçekleştirilmiştir . Her gösterge için (örneğin, sinapsların kapladığı mikrograf alanının yüzdesi), sağda ve solda PMVG olmak üzere iki ortalama kontrol değeri elde edildi. Bu ortalama değerler istatistiksel analize tabi tutuldu.
dikenler tarafından kaplanan ortalama alan bakımından tavukların hemisferleri ve grupları arasında fark yoktu ( Tablo 10.1). Aksispinöz ve aksodendritik sinapslar arasındaki ilişki de öğrenmeye bağlı olarak değişmemiştir. Ancak “eğitimsiz”
Tablo 10.1. SMVG'de sinaptik tomurcuklar ve dendritik dikenler tarafından işgal edilen alanın tahminleri (Bradley ve diğerleri, 1981)
Ortalama ve ortalama hata, eğitim türüne bağlı olarak ayrı ayrı gösterilir . ve yarımküre beyin. Her biri fotomikrograf temsil etmek Meydan beyin 80 um 2 .

Resim: 10.4. SMVG'deki sinapsın aktif bölgesinin postsinaptik sıkıştırmasının uzunluğu. Dikenlerdeki ve dendritik gövdelerdeki sinapslara ilişkin veriler birleştirilir. Farklı gruplar için ve yarım küreler için ayrı ayrı ortalama değerler verilmiştir (çift taramayla kaplı sütunlar - sol yarım küre; kısa çizgilerle kaplı sütunlar - sağ yarım küre). A. "Eğitimsiz" kuşlar. Sağ hemisferdeki ortalama postsinaptik kompaksiyon uzunluğu soldakinden %10.6 daha fazladır (P<0.025). B. "Yeniden eğitilmiş" kuşlar. Sağ yarıküre için ortalama sol yarıküreden 15 ± 16,4 nm daha az olmasına rağmen, fark (“az eğitimli” grupta gözlemlenenin tersi) istatistiksel olarak anlamlı değildir. (Bradley ve diğerleri, 1981.)
Kuşlarda, postsinaptik sıkıştırmanın uzunluğunda interhemisferik bir asimetri gözlendi (Şekil 10.4, K). Bu kuşlarda, sağ yarımkürede postsinaptik kompaksiyonun ortalama uzunluğu sol yarımkürede olduğundan önemli ölçüde daha uzundu (35±11.4 nm). "Yeniden eğitilmiş" tavuklarda gözlemlenen belirgin bir interhemisferik asimetri yoktu (Şekil 10.4,5). Aşırı eğitilmiş ve yetersiz eğitilmiş tavuklar için sol yarıkürelerdeki verilerin doğrudan karşılaştırılması, ek eğitimin postsinaptik sıkıştırma uzunluğunda ortalama 33 ± 14.6 nm, yani yaklaşık %10 (5<0,05) artışa yol açtığını gösterdi.
Ek eğitim , "yetersiz eğitim almış" kuşlarda bulunan hemisferik asimetriyi ortadan kaldırır. Stewart ve arkadaşları (1984) ayrıca kontrol tavuklarında eğitimle kaybolan bir asimetri buldular . Bu deneylerde tavuklarda pasif kaçınma geliştirildi (s. 143). Su ile nemlendirilmiş bir boncukta gagalanan kontrol tavukları ; deney grubuna civcivlerde iğrenme tepkisi uyandıran metil antranilat kaplı boncuklar verildi. 24 saat sonra , tavuklara kuru bir boncuk verildiğinde, kontrol tavukları onu gagalama eğilimindeyken, deney tavukları bundan kaçındı. Stewart ve meslektaşları ventral hiperstriatumdaki çeşitli sinaps parametrelerini ölçtüler. MVG'nin çalıştıkları alanı, görünüşe göre önündeki MVG'nin sınırları içinde yer alıyor (s. 92-95). Kontrol tavuklarında, postsinaptik sıkıştırmanın uzunluğunda asimetri gözlendi. Sol hemisferdeki ortalama uzunluk sağdakinden daha kısaydı. Asimetrinin ortadan kalkması muhtemelen sağ hemisferde postsinaptik kompaksiyonun uzunluğundaki bir azalmaya ve soldaki hafif bir artışa bağlıydı.
Damgalanma durumunda, interhemisferik asimetri , en azından kısmen, sol hemisferdeki postsinaptik kompaksiyonun uzunluğunu artırarak ortadan kaldırıldı . Bununla birlikte, öğrenmenin her iki yarımkürede de PMH'yi etkileme olasılığını göz ardı edemeyiz. Örneğin, sağ yarım küredeki değişiklikler çok hızlı gerçekleşebilir ve daha sonra, daha fazla öğrenmenin bir sonucu olarak, sol yarım küre onu "yakalar". Bu olasılığı araştırmak ve sinaptik değişiklikleri daha detaylı analiz etmek için çalışmalarımızı genişlettik: 1) 30 saatlik yaşamları boyunca karanlıkta tutulan 10 tavuktan oluşan bir grup ekleyerek; 2) “yetersiz” gruptaki civciv sayısını 9 kuş daha artırarak; 3) dendritik dikenler ve gövdelerde biten sinapsların bölünmesi (Şekil 10.3'teki örneğe bakın). Yeni bir partiden ilave civcivler alındığından , sonuçların bir havuzda toplanması için verilerin standardize edilmesi gerekliydi. Her iki çalışmada da bir grup "eğitimsiz" tavuk kullanıldı. Bu nedenle veriler bu gruplara göre standardize edilmiştir. Kolaylık sağlamak için, bu parametrenin öğrenme etkisi, karanlıkta tutulan kontrol grubundaki değeri ile ilişkilendirildi. İlk çalışma (Bradley ve diğerleri, 1981), damgalamanın presinaptik tomurcuklar ve dendritik dikenler alanı üzerinde bir etkisini ortaya koymadığından, bu parametreler ek tavuk gruplarında çalışılmamıştır (Hot ve diğerleri, 1985).
Hem aksospinöz hem de aksodendritik sinapslarla ilgili olarak, öğrenmenin, doku birim hacminde (sayı yoğunluğu) bulunan sinapsların sayısı üzerinde önemli bir etkisi bulunmadı. Eğitim ayrıca aksodendritik sinapsların postsinaptik sıkışmasının uzunluğunu da etkilemedi. Bununla birlikte , dikenli sinaps Sp L' nin postsinaptik yoğunluğunun genel ortalama uzunluğunu etkiledi . Bu değer yarım küre bağlı olarak değişir. LPMVH için , karanlıkta tutulan kuşlar için SPl değeri 286 ± 12,2 nm idi
.
Resim: 10.5. lPMSH'de dikenli sinapsların postsinaptik yoğunlaşmasının uzunluğu. Veriler, karanlıkta tutulan grupta 0 olarak alınan dikenli sinapsların postsinaptik yoğunluğunun uzunluğuna göre standartlaştırılmış ve ifade edilmiştir. Bu değer arasındaki ortalama farklar, "az öğrenilmiş" grup (çift taramalı sütun ) ve "aşırı eğitimli" grup (tek taramalı sütun) için göstergeler verilmiştir. Dikey çizgiler, bu ortalama değer ile 0 arasındaki farkın standart hatasını göstermektedir. “Aşırı eğitilmiş” tavuklar ile diğer iki grup arasındaki farklar anlamlıdır (P<0.01). (Noth ve diğerleri, 1985.)
Bu değer, 140 dakika eğitilmiş tavuklarda (49.2 ± 17.8 nm ile) önemli ölçüde daha yüksekti (Şekil 10.5). 20 dakikalık eğitim , "karanlık" grupla karşılaştırıldığında 8pb'yi önemli ölçüde etkilemedi . lPMCH'nin aksine, eğitimin etkisi altında pLMCH'de önemli bir değişiklik olmamıştır (Şekil 10.6).
Bu sonuçlar , (1) 20 dakikalık eğitime sinaps morfolojisinde önemli kaymaların eşlik etmediğini ; 2) daha fazla öğrenmenin önemli bir etkisi yalnızca lPMSH'de ortaya çıkar; 3) incelenen tüm parametrelerden, yalnızca dikenli sinapsların postsinaptik sıkıştırma uzunluğu öğrenmenin etkisi altında değişir. Karanlık kontrol ile karşılaştırıldığında, bir artış oldu
elli

Resim: 10.6. sPMSH'de dikenli sinapsların postsinaptik yoğunlaşma uzunluğu. Veriler standartlaştırılır ve Şekil 1'dekiyle aynı şekilde ifade edilir. 10.5. "Karanlık" gruptaki ortalama postsinaptik sıkıştırma uzunluğu 294 ± 7.2 nm idi. “Düşük eğitimli” ( çift hatlı sütun) ve “aşırı eğitimli” (tek kuluçkalı sütun) tavuklardaki alt değerler, bu değerden önemli ölçüde farklı değildir. (Noth ve diğerleri, 1985.)
ortalama uzunluk %17,2 arttı. Postsinaptik sıkıştırma yaklaşık dairesel bir şekle sahipse, çapı belirlenen uzunluğun basit bir türevidir (Weibel ve Bolender, 1973). Bu nedenle, uzunluktaki %17,2'lik bir artış, alanda yaklaşık %37'lik bir artışa karşılık gelmektedir.
Yukarıda açıklanan deneylerdeki civcivler, daha önceki deneylerde aynı uyaranla kullanıldıkları için tercih testine tabi tutulmadılar (Bateson, 1979). 21 dakika süreyle bir uyaran verilen tavuklar, bunu yeni bir nesneye tercih etmemiştir . Bu nedenle , “yetersiz eğitimli” tavuklarda tanıdık bir nesne tercihinin oluşmadığı varsayılabilir. Karanlık kontrol ile karşılaştırıldığında, sinapsların ve her iki PMVG'nin morfolojisinde herhangi bir değişiklik göstermediler. Buna karşılık, 81 dakika boyunca eğitilen civcivler, tanıdık uyaran için açık bir tercih gösterdiler (Bateson, 1979). Kullanılan " yeniden eğitilmiş" tavuk grubuna önemli ölçüde daha uzun bir süre boyunca bir uyarı verildi , bu nedenle karşılık gelen tercihi oluşturmaları beklenebilirdi. Ortaya çıkan morfolojik değişiklikler, hem yarımküreye hem de sinapsların tipine göre seçiciydi.
PMCH'nin öğrenmeye morfolojik yanıtında interhemisferik asimetrinin varlığına ilişkin veriler, bizi beyin hasarında damgalamada sol ve sağ PMCH'nin rolünü incelemeye sevk etti. Bu çalışmalar , hafızadaki bu beyin bölgelerinin işlevsel asimetrisi hakkında ek veriler sağladı (bkz. Bölüm 7).
Sinapsların yapısındaki değişikliklere ilişkin veriler, Sec.'de verilen elektrofizyolojik verilerle ilişkilendirilebilir. 10.1. Ancak bu iki deney serisinde farklı prosedürler ve nesneler kullanıldığı için bu ilişki oldukça zayıftır . Bu nedenle, dönen bir kırmızı kutu kullanılarak eğitilen civcivlerde morfolojik değişikliklerin gözlenip gözlenmediğini bulmaya karar verdik (Horn ve diğerleri, 1985).
Deneysel şema, Bölüm 1'de açıklananla aynıydı. 10.1. 21 saatlik yaştaki civcivler dönen bir tekerleğe yerleştirildi ve 2 saat boyunca kırmızı bir kutu ile sunuldu.Ertesi gün 1 saat eğitim tekrarlandı. Tekerlek dönüşlerinin sayısı, yaklaşma faaliyetinin bir göstergesi olarak hizmet etti. Eğitimden sonra civcivler karanlık bir inkübatöre geri gönderildi . Kırmızı kutuya yönelik güçlü bir tercihin oluştuğundan şüphe edilemeyeceği için tercih testi yapılmamıştır (bkz. Tablo 8.1). Civcivler 3 saat inkübatörde bekletildi, ardından yaklaşık 46 saat olduklarında anestezi ve perfüze edildi. Sonraki prosedür öncekiyle aynıydı, ancak elektrofizyolojik kaydın yapıldığı ek hiperstriatum alanından da doku blokları alındı (bkz. Bölüm 10.1). Her iki hemisferden bloklar alındı. Dikenler ve dendritik gövdeler üzerindeki sinapsların sayı yoğunluğu ve bu sinapslardaki postsinaptik sıkışmanın uzunluğu belirlendi.
Antrenmanın etkisi altında sadece bir ortalama puan önemli ölçüde değişti. Eğitimli tavuklarda lSMVG'deki dikenli sinapsların postsinaptik kalınlaşması, karanlık kontrolden önemli ölçüde daha uzundu (Şekil 10.7). Bu artışın derecesi , önceki deneyde "yeniden eğitilmiş" tavuklarda bulunandan önemli ölçüde farklı değildi . Bu nedenle, aynı tip sinapslar (dendritik dikenlerde ) ve beynin aynı tarafı (sol PMWG), yapay nesnelerle yapılan eğitimden etkilenmiştir, ancak bu iki çalışmada nesneler, deneysel tasarım ve tavukların yaşı perfüzyon süresi farklı olacaktır.
360
s
Resim: 10.7. lPMSH'de dikenli sinapsların postsinaptik yoğunlaşmasının uzunluğu. Kırmızı kutu kullanılarak 3 saat civciv eğitiminin etkisi . Kontrol "karanlık" grubu (gölgeli sütun) ve eğitimli tavuklar (siyah sütun) için ortalama postsinaptik sıkıştırma uzunluğunu gösterir . 16=2=6.97 nm'lik ortalama fark önemli ölçüde <P<0.03'tür). (Horn ve diğerleri, 1985.)
Bir elektrofizyolojik çalışmada, PMVG hücrelerinin kendiliğinden boşalma sıklığının, öğrenme sırasındaki yaklaşma etkinliği ile ilişkili olduğu bulundu (Şekil 10.1, A). lPMSH'de Sp L dikenli sinapsların postsinaptik yoğunluğunun uzunluğu ile yaklaşma aktivitesi arasında bir korelasyon var mı ? Analiz, antrenmanın 1. saatindeki yaklaşım göstergeleri ile lPMSH'de “Sp L ” arasında pozitif bir korelasyon ortaya çıkardı (r = 0.52; serbestlik derecesi sayısı 13; P <0.05). Eğitimin 2. ve 3. saatlerindeki yaklaşma tepkilerinin bağımlı bir göstergesi olarak kullanıldığında korelasyon anlamlı değildi (sırasıyla r = 0.12 ve r = 0.23 ). Korelasyon katsayısındaki bu düşüşün fonksiyonel önemi var mı? Kırmızı bir kutuya 20 dakika maruz kalmak bu nesne için bir tercih oluşturmak için yeterli olmasa da (Bateson, 1979), 1 saatlik eğitimden sonra yüksek tercih oranları elde edilebilir (bkz. Şekil 9.5). Bu oran, bu damgalama uyaranı kullanılarak 80 dakika (bkz. Şekil 9.7) veya 150 dakika (bkz . Böylece veriler, 1. saatteki öğrenme miktarının ve dolayısıyla ilişkili nöral değişikliklerin 2. ve 3. saatlerdekinden daha fazla olduğunu göstermektedir (bkz. Bölüm 3.2 ve Şekil 3.4).
Sp L'nin bireysel değerleri arasında korelasyon yok pPMCH veya ek hiperstriatum için anlamlı bir düzeye ulaşmadı.
Yorumlar ve akıl yürütme
Öğrenmenin sonuçları spesifik mi yoksa spesifik değil mi?
Postsinaptik sıkıştırmanın uzunluğundaki değişiklikler bellekle ilgili olmayabilir, ancak öğrenme prosedürünün başka bir yönünün sonucu olabilir. Örneğin, farklı tavuk gruplarında görsel deneyim veya lokomotor aktivite miktarındaki farklılıkları yansıtabilirler. Bu olasılıklar iki şekilde analiz edilebilir - bu çalışmanın verileri ve diğer deneylerin sonuçları temelinde. İlk veri dizisi göz önüne alındığında, öğrenme ek hiperstriatum ve pPMSH'deki herhangi bir göstergeyi etkilemediğinden , görsel deneyimin veya lokomotor aktivitenin sinapsların yapısı üzerindeki genel etkileri reddedilebilir. Karanlıkta tutulan grupta sinapsların gelişimi gecikebilir. Ancak bu durumda, beynin bölgesine, sol veya sağ yarım küreye ve sinapsların türüne özgü olduğundan, gelişimsel gecikme çok seçici olmalıdır . Bu tür lokomotor aktivite pek kritik bir faktör değildir. Kanıtlardan biri, yalnızca eğitimin 1. saatindeki yaklaşım faaliyetinin dikenli sinaptik temasın uzunluğu ile ilişkili olmasıdır . Eğitimin 2. ve 3. saatlerindeki yaklaşma etkinliği için karşılık gelen korelasyonlar istatistiksel olarak anlamlı bir düzeye ulaşmamaktadır . Ancak, eğitimin 1. saatindeki yaklaşım, kalan iki saattekinden daha zayıftır (Hot ve diğerleri, 1985). Bununla birlikte, PMVP'nin öğrenme sürecine katılımı hakkındaki iddia, yalnızca sinapslarındaki değişikliklerin öğrenmeden sonra tespit edildiği gerçeğine dayanıyorsa , genel itirazları çürütmek zor olacaktır . Ancak öyle değil. Önceki bölümlerde tartışılan morfolojik verilerin yanı sıra diğer veriler, PMVP'nin bellek süreçlerine dahil olduğuna işaret eder .
korelasyonlar
beyin homojenatlarından izole edilebilir (Matus ve Walters, 1975). Aracı reseptörlerin bu mühürlerde bulunduğunu gösteren güvenilir veriler vardır (Fagg ve Matus, 1984); bu nedenle, bu açıdan, postsinaptik yoğunlaşma , nöromüsküler bağlantıların postsinaptik zarlarına benzer (Cohen, 1980). Bu temelde, sıkıştırma uzunluğundaki bir artışın , postsinaptik zardaki reseptör sayısındaki bir artışla ilişkili olduğu varsayılabilir (Horn ve McCabe, 1987).
Yaklaşım üsleri ile Sp L arasındaki korelasyonların , bir yandan ve diğer yandan aynı göstergeler ile PMIG'deki kendiliğinden boşalma seviyesi arasında bir şekilde ilişkililer mi? Kedide lateral genikülat gövdeden görsel kortekse giden afferent liflerin büyük çoğunluğu dendritik dikenlerde son bulur (Colonnier ve Rossignol, 1969; Horning ve Garey, 1981); bu sinapsların doğası gereği uyarıcı olduğuna inanılmaktadır (Gray, 1959; Eccles, 1964). Japon bıldırcınlarında (Coturnix coturnix japonisa), görsel Wulst'a giden uzun afferent liflerin çoğu dendritik dikenlerde son bulur (Watanabe ve diğerleri, 1983). Sonuç olarak, dikenli sinapsların uyarıcı doğası hakkındaki varsayım , kuş beyninin nöronlarına kadar genişletilebilir . PMSG'deki sinapslarla ilgili bu varsayımı kabul edersek, beynin bu bölgesindeki yaklaşım aktivitesi ile morfofizyolojik değişiklikler arasındaki korelasyon için oldukça basit bir açıklama sunabiliriz. Civciv tanıdık uyarana doğru ne kadar uzun süre koşarsa, dikenli sinapsların postsinaptik sıkışmasının uzunluğu o kadar uzun oluyordu. Dikenlerde biten afferent liflerin, postsinaptik hücrelerin spontan impuls aktivitesinin oluşumunda rol oynadığını varsayalım. Postsinaptik sıkıştırma ne kadar uzun olursa , presinaptik lifteki spontan deşarjın postsinaptik hücrenin aktivitesi üzerindeki etkisi o kadar büyük olacaktır . Bu nedenle, presinaptik lifin aktivitesi sabit kabul edilse bile , sinaptik temas ne kadar uzun olursa, bu aktiviteyi postsinaptik nörona o kadar verimli bir şekilde iletecektir. Buna göre, yaklaşık aktivite ^Pl ile pozitif olarak ilişkiliyse , ayrıca spontan deşarjların ortalama sıklığı ile de pozitif korelasyon göstermelidir . Ancak bu korelasyon negatifti (Şekil 10.1, L). Böylece, tavuklar tanıdık nesneye yaklaştıkça Sp L o kadar yüksek , ancak nöronal deşarjların ortalama frekansı o kadar düşüktü . Değiştirilebilir sinapslara sahip hücreler kaydedilmekte olan hücreye göre inhibe edici ise , korelasyonun işaretindeki bu değişiklik meydana gelebilir .
10-3.3. Dendritik dikenlerin işlevleri
Dendritik dikenler ve modifikasyonları birçok araştırmacının ilgisini çekmiştir. Fareler 20 gün karanlıkta tutulduğunda birim başına düşen diken sayısı azalır.
140 uzunluğundadır (Valverde, 1967). Bu tür fareler ışığa maruz kaldıklarında , bu maruziyetin süresine bağlı olarak diken sayı yoğunluğu artmakta ve 4 gün sonra ışıkta büyüyen kontrol grubunda gözlenen seviyeden önemli ölçüde farklı olmayan bir düzeye ulaşmaktadır (Valverde, 1971) . . Fifkova ve van Harreveld (Fifkovâ ve Van Harreveld, 1977), sıçanların hipokampal bölgesindeki hücreler üzerindeki diken hacminin, onlara giden afferent yolların elektriksel uyarımı ile arttığını bulmuşlardır. Böyle bir değişikliğin fizyolojik sonuçları oldukça önemli olabilir (Rall ve Rinzell, 1973; Rall, 1978). Bazı dikenlerin boynu, sinaptik zarı içeren distal başlarından daha dardır (bkz. Şekil 10.3, K). Sinaps depolarize edildiğinde akım, sinapsın boynu boyunca ve dendritik gövde boyunca yayılır. Omurga boynunun çapındaki bir değişiklik , sinapsın verimliliğini veya "işlevsel ağırlığını" etkileyerek akımın dendritik gövdeye yayılmasını etkiler (Chang, 1952; Rall, 1974; Rall ve Rinzell, 1973; Koch ve Poggio , 1983). Mevcut yayılımdaki bu değişikliğin sonucu, bir nöronun sivri uçlu deşarj olasılığındaki bir değişiklik veya yerel sinaptik etkileşimin büyüklüğündeki bir değişiklik olmalıdır (örneğin , dendro-dendritik; bkz . Shepherd, 1979). Fifkova ve van Harreveld tarafından açıklanan omurga hacmindeki değişiklikler, omurga boyun çapındaki bir artışın varsayılan etkisiyle tutarlıdır , ancak böyle bir etkinin varlığına doğrudan kanıt sağlamaz . Boyun çapındaki küçük kaymalar gözden kaçmış olsa da, omurga hacminde herhangi bir değişiklik bulamadık (Bradley ve diğerleri, 1981).
Dendritik dikenler üzerine yapılan teorik çalışmaların çoğunda, araştırmacılar sinaps zarının pasif olduğu, yani bir aksiyon potansiyeli oluşturmadığı gerçeğinden yola çıkarlar. Eğer durum buysa, dikenli sinapsın etkinliği hiçbir zaman daha yüksek olamaz ve büyük olasılıkla dendritik sinapslarınkinden daha düşüktür ( Miller ve Jacobs tarafından yapılan incelemeye bakın, 1984). Bununla birlikte, eğer omurga zarı pasif değil aktifse, o zaman omurga uyarıcı postsinaptik potansiyeli (EPSP) artırarak güçlü bir sinaptik yükseltici görevi görebilir (Jack ve diğerleri, 1975; Miller ve Jacobs, 1984). Bilgisayar modellemesine dayanarak, aktif membranlı bir omurganın dendrite pasif membranlı bir omurgaya veya dendritik bir gövde üzerindeki sinapsa göre daha güçlü bir akım ilettiği öne sürülmüştür (Miller ve Jacobs, 1984). Bu nedenle, aktif zar dikenleri, bir nöronun ateşleme olasılığı üzerinde aksodendritik sinapslardan daha güçlü bir etkiye sahip olabilir. Bu düşüncelerden şu sonuç çıkıyor ki
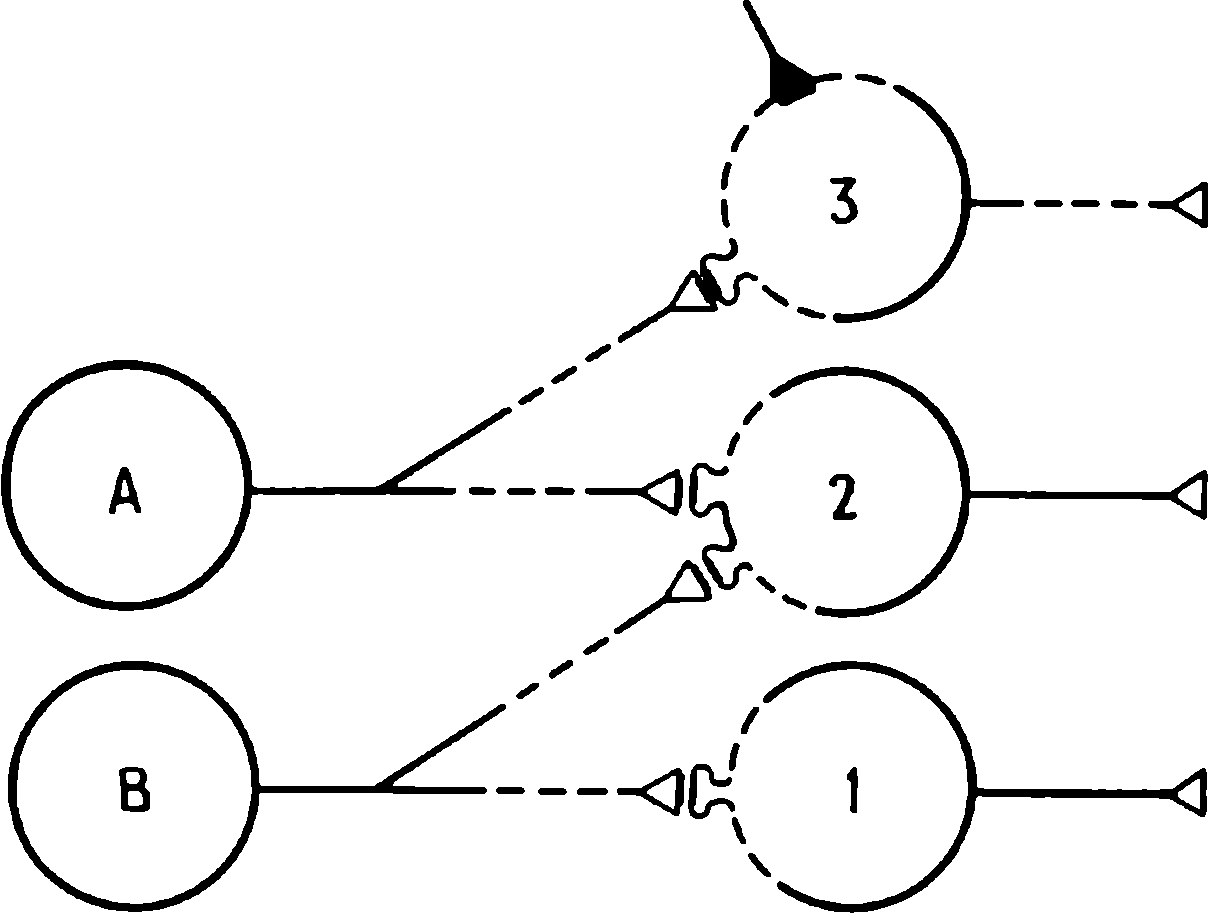
Resim: 10.8. Sinaps modifikasyon özgüllük modeli. Uyaran A lifini uyarır, ancak B lifini uyarmaz . Bu durumda, hücre 2'de bir deşarj meydana gelir. Hücre 3 , örneğin başka bir girdinin inhibitör etkisi altında olduğu için impuls oluşturma seviyesine kadar uyarılmaz. Postsinaptik yoğunluğun uzunluğunu artırmanın koşulu , pre- ve postsinaptik elemanların birbirine bağlı deşarjı ise, o zaman sadece A/2 sinapsı değiştirilecektir. Beyaz üçgenler uyarıcı sinapsları, siyah üçgenler ise inhibe edici sinapsları temsil eder. Kesik çizgiler ölçeklendirmeyi gösterir. Spinül başının çapı 0,5 µm'den biraz daha azdır; somanın çapı 50 kat daha büyük olabilir. Daha ayrıntılı açıklamalar için metne bakın.
postsinaptik sıkıştırmanın uzunluğundaki küçük değişiklikler , dendritik gövdelerdeki sinapslardaki temas bölgesinin uzunluğundaki bir değişiklikten ziyade, postsinaptik hücre deşarjlarını veya yerel sinaptik etkileşimleri düzenlemenin daha etkili bir yoludur.
Bir sinapsın bir postsinaptik hücre üzerindeki etkisi, kısmen hücre ile sivri ucun üretildiği bölge arasındaki mesafeye bağlı olabilir. Bu durumda, spike üretim bölgesinin daha etkin kontrolü, diğer şeyler eşit olmak üzere, uzaktaki sinapsları değiştirmek yerine bu bölgenin yakınındaki sinapsları değiştirerek elde edilecektir. Bu nedenle, birçok seçeneğin mümkün olduğu açıktır .
Sinaps Değiştirme Sınırlamaları
lifler tarafından aktive edilenlerle mi sınırlı yoksa bu değişikliğin postsinaptik hücrenin tüm sinapslarını kapsayan bir sınırlaması yok mu? Bu soru, sırayla, bir dizi soruyu gündeme getiriyor. Bunu göstermek için (Şekil 10.8), presinaptik lif A'nın kırmızı bir kutu gibi bir uyaranın sunulmasıyla etkinleştirilen liflerden biri olduğunu varsayalım. Fiber B, bu uyaran tarafından aktive edilmez. A ve B lifleri , sırasıyla 2-3 ve 1-2 hücrelerine göre presinaptiktir . A lifindeki bir impuls, hücre 2'de bir deşarja neden olur, ancak inhibisyon durumunda olan hücre 3'te değil. A lifindeki aktivite, postsinaptik yoğunluğun uzunluğunda bir değişikliğe neden oluyorsa , bu değişiklik A/2 temasıyla mı sınırlı yoksa bu uyaran tarafından etkinleştirilmeyen B/2'de de mi oluyor? Diğer bir olasılık da, A/3 de dahil olmak üzere A lifi tarafından oluşturulan tüm sinapsların, bu lifteki aktivite nedeniyle modifiye edilmiş olmasıdır. Sinapslardaki değişikliklerin özgüllüğü sorusu önemsiz değildir. Bir kedinin görsel korteksindeki tek bir nöron, 60.000'e kadar sinaptik terminal alabilir (Cragg, 1967) ve tek bir afferent lif , terminal dallarının dağıldığı yaklaşık 5.000 nöronu etkileyebilir (Sholl, 1956). Lifteki aktivite tek başına sinapsı değiştirmek için yeterliyse, bunun meydana gelmesi 5000 nöronun tümü ile bağlantıların değişmesine yol açacaktır. Ve eğer bu uyarılmış hücrelerden bazıları diğer hücrelere göre uyarıcı ise, o zaman bir nöronun boşalması sonucu beynin büyük bir bölümündeki sinapsların etkinliği artacaktır . Aynı zamanda, bir postsinaptik hücrenin aktivitesi, üzerinde sonlanan tüm sinapsların modifikasyonuna yol açıyorsa, bu, böyle bir hücrenin belirli bir sinir ağının oluşumuna katılma kabiliyetinde ciddi bir sınırlamaya yol açmalıdır.
Sinaptik değişikliklerin özgüllüğünü korumanın yollarından biri, yalnızca pre- ve postsinaptik öğelerin sıralarının çakıştığı sinapslarda morfolojik değişikliklere izin vermektir . Böyle bir varsayımın mantıksal sonuçları James (James, 1890) tarafından açıkça görülmüştür . Şöyle yazdı: "Eğer iki temel beyin süreci aynı anda aktifse veya birinin aktivitesi diğerinin aktivitesini hemen takip ediyorsa, o zaman biri ortaya çıktığında, daha sonra uyarılmanın diğerine yayılma eğilimi olacaktır" (James , 1890, cilt I, s. 566) . Hebb ( 1949, s. 62) bu görüşü paylaştı. Şöyle yazdı: "A aksonu B hücresine o kadar yakınsa, onu uyarabilir ve tekrar tekrar veya sürekli olarak deşarjlarına neden olabilir, o zaman hücrelerin birinde veya her ikisinde bir tür büyüme süreci veya metabolik değişiklik meydana gelir, böylece hücrenin yeteneği A hücresinin boşalmasına etkili bir şekilde neden olan B hücresi artar. Başka bir deyişle, özgüllük sorununun çözümü , sinapsların modifikasyonu, pre- ve postsinaptik elemanların aynı anda boşaltılmasıyla gerçekleşirse elde edilir. Bu nedenle, A lifi yalnızca 2. hücrede bir deşarja neden oluyorsa, o zaman yalnızca A / 2 sinapsı yükseltilir (Şekil 10.8), ancak B / 2 sinapsı veya A / 3 sinapsı değil, çünkü lif 2'nin deşarjı bir a ile ilişkili değildir. hücre 3'te deşarj (ayrıca bkz. bölüm 10.3.6).
elemanların birleşik deşarjının tekrar tekrar meydana gelmesi gerekliliğinin birçok avantajı vardır. Örneğin, hem pre- hem de postsinaptik nöronlar kendiliğinden aktif olabilir ve ateşlemeleri rastgele çakışabilir. Tekrarlanan eklem deşarjlarının gerekli olması , böyle rastgele bir olayın sonucu olarak sinaps modifikasyonunun meydana gelme olasılığını azaltır. Ek olarak , A lifi 2. hücrede bir ani artışa neden olursa, bu durumda bu boşalma bazen B lifinde kendiliğinden bir boşalma ile çakışacaktır. Lif B'deki ani yükselme 2. hücre için eşiğin altında olsa bile 2. hücrede bir boşalma meydana gelebilir. B lifindeki artış A lifindeki artışla sürekli olarak çakışmıyorsa, B/2 sinapsı değiştirilmeyecektir. Bu, B lifindeki artış hücre 2'deki artışla yeniden çakışmayacağı için gerçekleşecektir. Kuşkusuz, "tekrar" faktörü önemlidir: pre- ve postsinaptik elemanların ne sıklıkta birlikte boşaltılması gerektiği ve ne kadar süreyle boşaltılacağına bağlı olacaktır. fizyolojik özellikleri üzerinde bu sistem.
, sinaptik değişikliklerin yerel özgüllüğünü açıklar . Boşalmaların eşzamanlılığı , bu nedenle, değişiklikler için gerekli bir koşul olabilir . Hatta bazı sinapslar için eş zamanlı deşarjlar modifikasyon için yeterli bir koşul olabilir: eğer iki hücre birlikte yeterince ateşlenirse, aralarındaki bağlantı değişebilir (Barry ve Feher, 1981). Öğrenmeye dahil olan sinapslar için daha fazlasının gerekli olması mümkündür (bkz. Bölüm 6.4). Bu durumda, norepinefrin veya asetilkolin gibi ek bir faktörün etkisi gerekli olabilir (bkz. bölüm 9.3 ve s. 249). Gerçekten de, farklı nöronal sistemlerin onları değiştirmek için farklı maddelere ihtiyaç duyması mümkündür ; örneğin norepinefrin, sinapslarda değişikliklere neden olabilen büyük bir madde grubundan yalnızca biri olabilir .
Hücresel mekanizmalar
Postsinaptik sıkıştırmanın uzunluğundaki bir değişiklik, Dikenli kafaya yeni zar segmentlerinin eklenmesi veya sinaptik olmayan zarın modifikasyonu ve değiştirilmesi nedeniyle olabilir . İlk varyantta, Shipik'in hacminde bir artış beklenmelidir. Bu olmaz (Tablo 10.1); bu nedenle, zarın yeni, ek bölümlerinin omurganın başına dahil edilmesiyle sinaptik mührün uzunluğunda bir artış elde edilemez. Eğer böyle bir kaynaşma gerçekleşmişse, buna , sinaptik temasın dışında karşılık gelen zar alanının ortadan kaldırılması eşlik etmelidir . Aksi takdirde, bu zar dönüştürülmeli ve sinaptik mührün içine dahil edilmelidir .
Kimyasal sinapslarda, nörotransmiterler postsinaptik zardaki reseptör proteinlerine bağlanarak etki ederler . Bir proteine bağlanan moleküle ligand evi denir. Bazı sinapslarda nörotransmitter, zarın iyon kanallarına bağlanır ve bunların konformasyonunu değiştirir. Ligand-kapılı iyon kanalları açılır ve zar boyunca ortaya çıkan iyon akışı, zar potansiyelinde bir kaymaya yol açar. Norepinefrini bağlayanlar da dahil olmak üzere diğer sinapslarda, bu nörotransmitter, postsinaptik hücreyi iyon kanallarını açarak değil , "ikinci haberci" aracılığıyla etkileyebilir (bkz . Kehoe ve Marty, 1980; Hartzell, 1981). Bu sinapslarda, nörotransmitter - "ilk haberci" - zar reseptörlerine bağlanır ve böylece hücre içinde bir olaylar zinciri başlatır. Bu olayların spesifik doğası bağlı reseptöre bağlıdır. Örneğin, norepinefrin [31-adrenerjik reseptörlere] bağlandığında, adenilat siklaz enzimi aktive olur ve “ikinci haberci” olan siklik adenoz monofosfat (cAMP) konsantrasyonu artar. cAMP, protein kinazı aktive ederek hücredeki birçok işlemi kontrol eder. Bununla birlikte, norepinefrin, ainoradrenerjik reseptörlere bağlanırsa , sitozolde serbest kalsiyum seviyesinde bir artışa yol açabilen bir işlemler zinciri oluşur (aşağıya bakın). Kalsiyum ayrıca ikinci bir haberci olarak da hareket edebilir.
, kalsiyum bağlayıcı proteinlerin yapısını değiştirerek gerçekleştirebilir (Kretsinger, 1981). Bu proteinlerden biri olan kalmodulin, incelenen tüm hücrelerde mevcuttur. Siklik AMP gibi, kalmodulin de hücredeki çeşitli süreçleri düzenler ve bunu birçok proteinle etkileşerek yapar. Bazı durumlarda, ikinci arabulucular hücre tepkisinin düzenlenmesinde ortaklaşa yer alabilirler ve etkileri sadece işbirlikçi değil, aynı zamanda karşıt da olabilir (Rasmussen, 1980). Çalışmalar , kalsiyum iyonlarının bir aksiyon potansiyeli meydana geldiğinde voltaja bağlı kanallardan geçerek hücreye girebileceğini göstermiştir (incelemelere bakın, Adams ve diğerleri, 1980; Neering ve McBurney, 1984). Llinas ve diğerleri (Llinâs ve Sugimori, 1980; Llinâs ve Yarom, 1981) , kalsiyumun bazı memeli merkezi nöronlarının dendritlerine girdiğine dair net kanıtlar elde ettiler. Norepinefrine ek olarak birkaç ligandın, hücre organellerinde biriken kalsiyumu harekete geçirerek hücre içi kalsiyum seviyesinde bir artışı uyarabildiği de gösterilmiştir (Berrige, 1986). Düz endoplazmik retikulumun zarlarının kalsiyum biriktirdiği bilinmektedir (McGraw ve diğerleri, 1980; Rasmussen ve Barrett, 1984). Dendritlerin sitoplazmasında bulunan düz endoplazmik retikulumun büyümeleri dendritik dikenlere ulaşabilir (Peters ve Kaiserman- Abramof , 1970). Boyundan spinülün başına geçen bu tübüler sistem , spinöz aparatı oluşturmak için bir zarlı yapı kompleksi ile birleşir (Gray, 1959; Gray Guillery, 1963; Westrum ve diğerleri, 1980) . Hücrede tipik bir dikenli aparat her zaman mevcut değildir , ancak dendritik dikenlerde genellikle düz zarlarla çevrili basit vakuoller görülür (Peters ve diğerleri, 1976). Önerilen modelde, bu zarların kalsiyumu tuttukları varsayılmaktadır.
tarif edilen kaymalara yol açan hangi moleküler değişikliklerin iki gereksinimi karşıladığı düşünülebilir : 1) pre- ve postsinaptik hücrelerin ortak deşarj ihtiyacı; 2) sinaptik değişikliklerin belirli bir aktif maddenin varlığına bağımlılığı? Kalsiyum iyonlarının doğrudan veya kalmodulin aracılığıyla, postsinaptik sıkıştırma yapısındaki değişikliklerin altında yatan süreçleri etkilemesi mümkündür. Eklem deşarjlarının gerekliliği, pre- ve postsinaptik nöronlarda yakın zamanda aksiyon potansiyellerinin tekrar tekrar ortaya çıkmasıyla karşılanır . Presinaptik fiberin postsinaptik hücreyi uyarması beklendiğinden, sinaps uyarıcı olmalıdır . Bu, SMVG'deki dikenli sinapsların değiştirilebileceğine dair verilerle desteklenir (Bölüm 10.2 ve 10.3.3). Akson terminalinde bir dürtü belirdiğinde, nörotransmiter serbest bırakılır. Muhtemelen postsinaptik sıkıştırmada bulunan reseptör moleküllerine bağlanır . Ligand kapılı iyon kanalları açılır, akım zardan akar ve EPSP şeklinde kaydedilir. Genliği küçükse, postsinaptik hücrenin zarı hızla dinlenme durumuna döner. EPSP yeterince büyükse, postsinaptik hücrede akımın dendritler boyunca yayılmasına neden olan bir aksiyon potansiyeli üretilir. Bu nedenle, temasın postsinaptik kısmı için EPSP, presinaptik lifte bir artış anlamına gelir ve bir aksiyon potansiyeli, postsinaptik bir nöronda bir artış anlamına gelir. Dolayısıyla bu olaylar, pre- ve postsinaptik elementlerin birleşik deşarjının "tanınmasının" temelidir. Aşağıdaki akıl yürütmede, dendritik omurganın zarının pasif olduğu varsayılmaktadır (s. 241-243). Bununla birlikte, genel olarak, bu argümanlar, aktif bir zara sahip sivri ucun varyantına da genişletilebilir . Bu aktivite EPSP ile yakından ilgili olacaktır; aksondaki sivri uç daha sonra oluşacak ve postsinaptik hücre tarafından üretilen gerekli sinyali oluşturacaktır.
Ortak uyarımı temsil eden bu iki "sinyal", postsinaptik sıkıştırmada bir değişikliğe yol açan bir dizi olayı tetiklemek için kullanılır . Aşağıdaki fenomen zinciri önerilebilir, ancak bu şemanın çoğu spekülatif akıl yürütmeye dayanacaktır. Presinaptik terminal tarafından salınan aracının hücre içine kalsiyum akışına neden olduğu (MacDermott ve diğerleri, 1986) veya dendritik omurganın hücre içi oluşumlarında biriken kalsiyumu harekete geçirdiği varsayılmaktadır . EPSP'lerin etkisi altında postsinaptik hücrede bir aksiyon potansiyeli üretilirse, voltaj kapılı kalsiyum kanallarından daha fazla kalsiyum akışı , postsinaptik yoğunlaşma yakınında omurga başındaki bu iyonun yerel konsantrasyonunu artırabilir. Bu yüksek kalsiyum konsantrasyonunun ancak arabulucunun etkisiyle ve voltaja bağlı kanalların açılmasıyla ilişkili kalsiyum iyonlarının hareketinin yakın bir zaman dizisinde gerçekleşmesi ve bu olayların tekrarlanması durumunda yaratıldığı varsayılmaktadır (s. 244). . Ulaşılan kalsiyum konsantrasyonu, modifikasyonun yalnızca iki sinyalin çakışmasını gerektirdiği sinapslarda kalmodulin'i aktive etmek için yeterli olabilir. Ek olarak özel bir maddenin varlığının gerekli olduğu sinapslarda , kalsiyum konsantrasyonundaki bu artış, gerekli olduğu halde, kalsiyum bağlayıcı proteini aktive etmek için yeterli olmayabilir. Calmodulin, dört kalsiyum bağlama lokusuna sahiptir; molekülü aktive etmek için en az ikisinin kalsiyum iyonları tarafından işgal edilmesi gerekir . Adrenerjik reseptörlere bağlanan norepinefrin varlığında ek kalsiyum mobilizasyonu meydana gelebilir. Bu iyonun yerel konsantrasyonu, kalmodulin konformasyonunu değiştirmek için yeterli olabilir. Sinaptik membran reseptörlerinin fosforilasyonunda yer alan bir enzimi aktive eder (Smilowitz ve diğerleri, 1981). Merakla, kalmodulin bağımlı protein kinazın yanı sıra kalmodulin (Grab ve diğerleri, 1980; Kennedy ve diğerleri, 1983; Kelly ve diğerleri, 1984) postsinaptik kalınlaşmada saptanır. Kalsiyumun etkisi çok lokal olabilir, çünkü sitozoldeki serbest kalsiyum hızla iç zarlar tarafından yakalanır (Blaustein ve diğerleri, 1980).
Baskılama için belirli konsantrasyonlarda norepinefrin gereklidir, ancak sinaps modifikasyonunda yer alan tek madde norepinefrin değildir. Damgalamanın şiddeti, lokomotor aktiviteye, uyanıklık düzeyine ve dikkatine bağlı olabilir ve bu süreçlere, çeşitli aracıları sentezleyen çeşitli nöronal substratlar aracılık edebilir. Örneğin , daha hızlı etki eden bir aktivasyon sisteminin etkisinin, noradrenerjik sistem için beklenen etki ile özetlenmesi mümkündür. Memelilerin medial septal çekirdeğinin uyanma reaksiyonunun meydana gelmesinde rol oynadığına inanılmaktadır (Green ve Arduini, 1954). Civcivde medial septal çekirdek SMVG'ye bağlantılar gönderir ve kolinerjik görünmektedir (Davis & Noth, 1983). PMVG'de muskarinik kolinerjik reseptörler vardır (Bradley ve Horn, 1981). Bu tür reseptörler, aracı bağlanması üzerine, hücre zarlarında bulunan inositol fosfolipidleri hidrolize edebilir (Berridge, 1986; Berridge, Irvine, 1984). Hidroliz, muhtemelen hücre içi depolardan kalsiyumu harekete geçirerek, hücredeki serbest kalsiyum seviyesinde bir artışa yol açar . Bu düşüncelerden, sitoplazmada serbest kalsiyum seviyesini etkileyen ve böylece kalmodulin aktivasyonunun derecesini düzenleyen birçok faktörün işbirliğine dayalı bir etkileşiminin mümkün olduğu görülebilir (Bear ve Singer, 1986). Böylece plastisite, nöronların moleküler mekanizmaları tarafından düzenlenebilir (Berridge, 1984). Bu tür mekanizmalar aracılığıyla, örneğin kalmodulin aktivasyonunda doğrusal olmayan bir artış elde edilebilir. Doğrusal olmayan etkileşimler, bir izin kaydı için bir sinyal görevi gören ikinci aracının ya hep ya hiç reaksiyonuna yol açabilir. Bu sinyal, elektriksel dürtü aktivitesini hızlı bir şekilde morfolojik bir değişikliğe dönüştürebilir ve böylece daha kararlı bir hafıza temeli oluşturabilir.
Reseptörler sürekli olarak değiştirildiğinden, muhtemelen dendritik taşımayı içeren hücre gövdesi tarafından sürekli olarak yeniden sentezlenmeleri gerekir (Fambrough, 1981). PMVG'de damgalama , postsinaptik mührün ortalama uzunluğundaki ve sonuç olarak omurga zarında kapladığı alanda bir artışla ilişkilidir . Bu, reseptör proteinin alımını arttırırsa, protein ve RNA sentezi artmalıdır. Belki de damgalamanın biyokimyasal çalışmalarında ortaya çıkan bu artıştı (bölüm 2 ve 3). Eğitimden sonra , sinaptik yoğunlaşmadaki reseptör sayısı eskisinden daha fazladır (Horn ve McCabe, 1987); bu nedenle, hücre gövdesinde reseptör proteininin sentezinde sürekli bir artış meydana gelmelidir. Nöromüsküler kavşak için gösterildiği gibi reseptör devir hızı yüksek değilse (Changeux ve Mikoshiba, 1978; Fambrough, 1981), o zaman protein sentezinde sadece hafif bir artış meydana gelir. Bu , eğitimden sonraki gün ölçülen radyoaktif urasilin RNA'ya dahil edilmesinin eğitim süresinin süresine bağlı olarak önemli ölçüde değişmediğini gösteren verilerimizi açıklayabilir (Şekil 3.6.5).
Pre- ve postsinaptik değişiklikler
sinaptik temasın post-sinaptik kısmına odaklandı . Bu tamamen ampirik bir temelde yapıldı. Sinapstaki her iki karşıt zar da elektron yoğun malzeme içerir. Bazı sinapslarda bu mühürler asimetriktir ve postsinaptik tarafta yer alan daha belirgindir. Diğer sinapslarda mühürler simetriktir (Gray, 1959; Colonnier, 1968). Postsinaptik zar en sürekli olarak belirgin bir kalınlaşmaya sahip olduğundan, PMSG ve aksesuar hiperstriatum çalışmasında bir nesne olarak seçilen bu zardı (Bradley ve diğerleri, 1981; Horn ve diğerleri, 1985).
Sinapsın hangi tarafının plastik değişikliklerin bölgesi olabileceği sorusu geçmişte defalarca tartışılmıştır . Hebb ( 1949, s. 62) şöyle yazmıştı: "Bir hücrenin diğerinde bir deşarj üretme yeteneğinin geliştirilmesine ilişkin en bariz ve umarım en olası varsayım, sinaptik tomurcukların ortaya çıkması ve böylece alanın artmasıdır. Afferent akson ile götüren hücrenin gövdesi arasındaki temas ”. Griffith ( 1966-1968) benzer bir görüş benimsedi ve birlikte uyarım gereksinimini karşılayan bir bağlantı değiştirme mekanizması önerdi. Bu yazarın görüşlerine göre, sinapsların özgüllüğü, presinaptik terminal üzerinde karşılıklı bir etkiye sahip olan postsinaptik eleman üzerinde bir maddenin salınmasıyla sağlanabilir . Aslında, presinaptik mekanizmanın plastik değişikliklerin düzenlenmesine dahil olduğuna dair bazı deneysel veriler vardır. Bu, sifon retraksiyonunun klasik savunma refleksinin gelişimi sırasında deniz yumuşakçası Aplysia'da gösterilmiştir . Veriler, uyarıcı bir akım geçirmek için bir duyu nöronuna bir mikroelektrodun yerleştirildiği bir çalışmada elde edildi (Hawkins ve diğerleri, 1983). Bu nöronun aksiyon potansiyelleri, kuyruk sinirine uygulanan bir dizi elektriksel impuls olan koşulsuz bir uyaranla birleştirildi. Sifon kasını kontrol eden nörona bir mikroelektrot da yerleştirildi . Bu nöron, kaydedilmekte olan duyusal nörondan bir giriş sinyali aldı . Duyusal nöronun spike deşarjı koşulsuz uyarandan önce geldiğinde, duyusal sinyalin motor nöron üzerindeki etkisi kolaylaştırılmıştır. Bu rahatlama presinaptik gibi görünmektedir (ayrıca bkz. Walters ve Bryne, 1983).
Bu durumda kabartma, Hebb'in pre- ve postsinaptik elementlerin birlikte boşaltılması gerekliliği hakkındaki kuralına uymadı (Bölüm 10.3.4). Daha sonra Carew ve diğerleri (1984), benzer bir deneysel tasarım kullanarak (Hawkins ve diğerleri, 1983), motor nöron aktivitesinin kolaylaştırma için ne gerekli ne de yeterli olduğunu buldular. Duyusal nöronu ve postsinaptik motor nöronu doğrudan elektriksel stimülasyonla uyararak, duyu nöronunun boşalması motor nöronun boşalmasından hemen önce gelecek şekilde organize ettiler. Bu prosedür, duyusal ve motor nöronlar arasındaki temasta iletimi kolaylaştırmadı. Ancak bu sonuç, öğrenmede yer alan sinapslar için beklenmeyen bir sonuç değildir. Griffith ( 1966, 1967, s. 34), Hebbian sinapsındaki iletim etkinliğindeki değişikliklerin yalnızca takviye ile ilişkili bazı ek faktörlerin varlığında meydana gelebileceğine işaret etti (ayrıca bkz. bölüm 6.4 ve 9.3), yani eklem sinaps öncesi ve sonrası elemanların uyarılması, sinapsı değiştirmek için yeterli değildir . Sinaps kolaylaştırma sırasında böyle bir aktiviteye ihtiyaç olup olmadığı sorusunu aydınlatmak için , Carew ve diğerleri ( 1984) bir motor nöronu "öğrenme" prosedürü sırasında aksiyon potansiyelleri üretmeyecek şekilde hiperpolarize ettiler . Koşulsuz bir uyaranın eşlik ettiği bir elektriksel uyaranla bir duyusal nöronun uyarılmasından oluşuyordu . Duyusal nörondan motor nörona iletim kolaylaştırıldı, bu da bu kolaylaştırmaya bir tür "güçlendirici" faktörün katıldığını gösterdi. Elde edilen sonuçlardan , sinaptik iletimi kolaylaştırmak için pre- ve postsinaptik elemanların ortak deşarjının gerekli olmadığı da anlaşılmıştır .
Bu çalışmalarda (Hawkins ve diğerleri, 1983; Walters ve Brunet, 1983) , koşulsuz uyaranın , duyusal nöronların terminalleriyle dallanmış temasları olan bir grup nöron aracılığıyla etki ettiği varsayılmıştır . Eğer bu böyleyse ve kolaylaştırma gerçekten de presinaptik bir yapıya sahipse, o zaman kolaylaştırma duyusal nöronların tüm terminallerinde gerçekleşmelidir . Bir yandan, kolaylaştırma sifonu kontrol eden motor nöronlarda son bulan terminallerle sınırlıysa, o zaman işlevsel etki spesifik olacaktır , ancak onu oluşturmak için Hebbian sinapslarının varlığı gerekli değildir . Dahası, sensorimotor refleksin "lehimlenmiş" yayındaki bu tür iletim duyarlılaşmasının herhangi bir özel şekilde öğrenme ile ilgili olup olmadığı şüphelidir . Öte yandan, bir duyu nöronu, kaçınma refleksinde yer almayan kasları innerve eden motor nöronlarda da sinaps yaparsa ve tüm bu sinapslar rahatlarsa, belirli bir davranışsal etkinin nasıl sürdürülebileceğini anlamak zordur (Bölüm 10.3.4). Bununla birlikte, Hebb kuralının değiştirilmesine izin verilirse, özgüllük korunabilir. Kayıt yapılan motor nöron, sifon innerve eden kaslardan biridir (Kupfermann ve ark., 1974). Bu nöronlar işlevsel bir grup oluşturabilir, böylece bir duyu nöronu ateşlendiğinde tüm motor nöronlar ateşlenir. Bir motor nöronun hiperpolarizasyonla etkisizleştirilmesi , geri kalanının boşalmasını engellemeyecektir. Değiştirilmiş sinaps bu deşarj hakkında bir sinyal alırsa , fonksiyonel motonöron grubunun ateşlenmesi hakkında bilgi iletecektir . Bu nedenle, eğitim sırasında, Hebb ve Griffith tarafından öne sürülen genel tipteki girdi, çıktı ve pekiştirici sinyallerin bir kombinasyonu olacaktır. Bununla birlikte, Hebb'in (Hebb, 1949, s. 61), kendi hipotezini göz önünde bulundurarak, " yetişkin hayvanlarda bu şekilde doğrudan duyu-motor bağlantılarının oluşmasının söz konusu olmadığını" belirttiği belirtilmelidir.
kritik bir değişikliğin , postsinaptik öğede meydana gelme olasılığı daha önce tarafımızdan tartışılmıştır:
, etkinleştirildiğinde zarın daha geniş bir alanını depolarize edebilen ve daha büyük olasılıkla post- sinaptik bir nöronun ateşlenmesine neden olabilen sinaptik tomurcukların boyutunda bir artış olabilir. Bununla birlikte, yazarın bilgisine göre, böyle bir büyüme hiç gözlemlenmemiştir.Nöronlar arasında işlevsel bir bağlantının kurulmasına yol açan alternatif bir varsayımsal değişiklik bölgesi, postsinaptik zar olabilir. Birkaç eğitimli tahminde bulunursak, sinapsın bu kısmındaki değişikliklerin iletimi etkilemesi muhtemeldir. Sinaptik tomurcuklardan salınan nörotransmitter moleküllerin sayısı, bu moleküllerin postsinaptik zarda bağlandığı reseptörlerin sayısını aşarsa, bu tür reaktif grupların sayısındaki artış, sinaptik iletimin etkinliğinde bir artışa yol açmalıdır. Reseptör moleküller muhtemelen bir protein veya lipoprotein olduğundan, sentezleri büyük olasılıkla hücre sitoplazmasındaki ribonükleik asit tarafından kontrol edilmelidir ” (Horn, 1962, s. 276) ►
Bu yazıldığından beri, diğer yazarlar sinaps fonksiyonlarındaki değişikliklerin postsinaptik lokalizasyonunu ileri sürmüşlerdir ( Stent, 1973; Heidman ve Changeux, 1982); PMIG çalışmasının sonuçları ayrıca postsinaptik yapının rolüne işaret ediyor . Ancak bu çalışmalar , bazı presinaptik yapılardaki değişiklikleri dışlamaz . Gerçekten de, sinapsın yalnızca bir tarafının değiştirilebileceğine inanmak için hiçbir neden yoktur . Kosover (Kosower, 1972), birincil değişikliğin temasın presinaptik tarafında meydana geldiğine inanıyordu. Arabulucu içeren veziküllerin salındığı presinaptik membran alanında bir artış önerdi . Kosower, sinaptik terminalden aracı çıkışındaki bir artışın "alıcı sayısında bir artışa ve postsinaptik reseptör bölgesinde bir artışa yol açacağına" inanıyordu (Kosower, 1972, s. 3295). Kısacası, bir sinapsın bir tarafındaki değişiklik diğer tarafında yankılanabilir. Verilerin gösterdiği gibi, ilk değişiklik postsinaptik ise, o zaman postsinaptik zardaki bir reseptöre ligand bağlanmasıyla tetiklenen müteakip kimyasal olaylar zinciri , presinaptik terminal üzerinde geri dönüşümlü bir etkiye sahip olan yerel bir nörohormonun sentezine yol açabilir . Örneğin, asetilkolin bir membran reseptörüne bağlandığında, inositol lipid, inositol trifosfat ve diasilgliserole hidrolize olur . İkinci bir aracı olarak diasilgliserol, araşidonik asit yoluyla prostaglandin hormonunun sentezini etkileyebilir (Berridge, 1982; Berridge, Irvine, 1984; Nishizuka, 1984). Temasların post- ve presinaptik bileşenleri arasında benzer bir etkileşim hipokampta bulundu. Belirli koşullar altında, bu yapıda sinaptik iletimin uzun vadeli güçlenmesi (LTP) indüklenebilir. LTP'nin başlatılması , postsinaptik süreçlere bağlı gibi görünmektedir (Collingridge ve diğerleri, 1983) ve LTP'nin sürdürülmesi , muhtemelen presinaptik terminallerden aracı amino asitlerin salınmasındaki değişikliklerle belirlenir ( Lynch ve diğerleri, 1985; Bliss ve diğerleri. ., 1986).
, özellikle dikenli sinapsların etkinliğindeki bir değişikliğin beklenen bir sonucu, presinaptik bir nörondaki bir artışın, bir post sinaptik nöronda bir artışa veya bir postsinaptik hücrenin komşu hücrelerle etkileşiminde bir değişikliğe neden olma olasılığındaki artıştır. nöronlar. Sinapsları bu şekilde değiştirilen nöronlar, eğitimden sonra eskisinden daha sık birlikte ateşlenir. Böylece öğrenme sinir sisteminde bir "iz" bırakır. Bu iz, başlangıçta bir nesnenin sunumu üzerine etkinleştirilen bir grup nöronda sabitlenirse, bu grup onun nöral temsilini oluşturacaktır (Hebb, 1949; Milner, 1957). Bu nöron grubundaki kararlı , kolaylaştırılmış iletim böylece bu nesnenin ezberlenmesi için temel oluşturacaktır ; bellek izinin bağlamsal zenginliği, bu grubun diğerleriyle eşzamanlı olarak oluşturulan ve bir zaman damgasıyla "işaretlenen" bağlantıları ile sağlanabilir. Ancak izin bu şekilde mi oluşturulduğu, bir hologramın nöral analoğu (Pribram, 1966; Longuet-Higgins, 1968) veya başka bir şekilde mi işlev gördüğü bilinmiyor (bkz. tartışma, Kohonen, 1984). .
İşlevsel Etkiler
Tavuklarda herhangi bir görsel deneyim olmasa bile doldurulmuş tavuk tercihi gelişebilir. Bu tercih, civcivlerin bir tür uyarana maruz kalması koşuluyla zamanla gelişir (Bölüm 8.2). Bu bariz tercihten sorumlu nöral devrelerin işlevsel olarak dolaşımdaki testosteron seviyelerine bağlı olabileceği öne sürülmüştür . Bu varsayımın dayandığı kanıtlar ikinci derecedendir, ancak testosteronun böyle bir sistemi nasıl etkileyebileceği sorusunu gündeme getirir (Bölüm 9.6). Bir olasılık, bu farazi sistemin nöronları arasındaki sinapsların yumurtadan çıkma sırasında ve sonrasındaki birkaç saat boyunca işlevsiz olmasıdır. Belirli koşullar altında , bir süre sonra tavuk dolması tercihi ortaya çıkıyor. Görünüşe göre, bu durumlarda sistemin nöronları arasında sinaptik iletim kurulur ve bu testosteronun katılımıyla gerçekleşir. Steroid bir hormon olan testosteronun, örneğin norepinefrin gibi hedef hücreler üzerinde etki etmediğine inanılmaktadır. Bu steroid hormon, sitozoldeki bir reseptöre bağlanır . Aktive edilmiş reseptör daha sonra, spesifik bir proteinin sentezine yol açan bir işlemler zinciri dahil olmak üzere kromatine bağlanır (Yamamoto ve Alberts, 1976). Beynin birçok bölgesindeki nöronlar seçici olarak steroid hormonları yakalar (Goy ve McEwen, 1980). Özellikle ilginç bir örnek, penis reflekslerini kontrol eden devrenin bir parçasını oluşturan sıçan omuriliğinin alt kısmındaki belirli nöronlar tarafından sağlanır . Bu nöronlar testosteronu depolar. Arnold ( 1981), androjenlerin bu motor nöronlar üzerinde hareket ettiğini ve böylece çiftleşme davranışından sorumlu devreleri etkilediğini ileri sürer. Hormonun bu hücreler üzerinde tam olarak nasıl etki ettiği hala bilinmiyor. Bu , yukarıda açıklandığı gibi kromatin üzerindeki bir etki yoluyla meydana gelebilir . Bununla birlikte, hormonun sinir hücreleri üzerinde daha doğrudan bir etki olasılığı göz ardı edilemez, çünkü uygulandıktan birkaç dakika ve hatta saniyeler sonra elektriksel aktivitelerini değiştirebilir (Andrew, 1983).
Özet
sunularak eğitildiklerinde, SMVG'deki nöronların aktivitesinde bir değişiklik tespit etmek mümkün oldu . Nöronların spontan aktivitesi, eğitim sırasındaki yaklaşma aktivitesi ile negatif olarak ilişkilidir: kutuya ne kadar çok tavuk yaklaşırsa, nöronal deşarjların sıklığı o kadar düşüktü. Bu etki, aksesuar hiperstriatumda saptanmadığı için bölgesel olarak spesifikti . Doldurulmuş tavukla eğitilmiş civcivlerde anlamlı bir korelasyon bulunmadığından, aynı zamanda uyarana özeldi. Kırmızı bir kutu veya yatay olarak konumlandırılmış yanıp sönen bir dikdörtgen kullanılarak eğitilen tavuklarda , PMVG sinapslarının yapısındaki değişiklikler ortaya çıktı. Her iki durumda da, öğrenme , dikenli sinapsların postsinaptik sıkışmasının uzunluğundaki bir artışla ilişkilendirildi. Sinapslardaki değişiklikler sol PMVG'de gözlendi, ancak sağda gözlenmedi. sPMSH'deki dikenli sinapsların postsinaptik yoğunluğunun uzunluğu, öğrenme sırasındaki yaklaşımın etkinliği ile ilişkilidir. Sinapsların yapısındaki değişikliklerin olası hücresel mekanizmaları ele alınmıştır. Dendritik dikenlerdeki değişikliklerin önemi, bu oluşumların nöronal aktivitenin düzenlenmesi ve hafıza süreçlerindeki olası işlevi ile bağlantılı olarak tartışılmaktadır .
EDEBİYAT
Adams DJ, Smith SJ, Thomson SH Yumuşakça somasında Ionie akımları, A. Rev. Neurosci., 1960, 3, 141-167.
Andrew RJ Östradiol ve testosteronun genç evcil civcivde dikkat dağınıklığı ve hafıza oluşumu üzerindeki spesifik kısa vadeli gecikme etkileri. İçinde: Yüksek omurgalılarda hormonlar ve davranış (eds. J. Balthazart, E. Prove ve R. Gilles), 1983, s. 461-473, Springer, W. Berlin.
Arnold AP Omurgalı davranışının kontrolünde steroid hormon etkisinin mantıksal seviyeleri, Am. Zool., 1981, 21, 233-242.
Baranyi A., Feher 0. Sinaptik kolaylaştırma, neokortekste yakınsak yolların eşleştirilmiş aktivasyonunu gerektirir , Nature, Londra., 1981, 290, 413-415.
Bateson P. Civcivlerde damgalama sırasında yeni bir uyarana kısa süre maruz kalma ve bunun sonraki tercihler üzerindeki etkisi, Anim. Leam. Behav., 1979, 7, 259-262.
Ayı MF, Şarkıcı 1F. Görsel kortikal plastisitenin asetilkolin ve noradrenalin ile modülasyonu, Nature, Londra., 1986, 320, 172-176.
Berridge MJ Hücre salgısının düzenlenmesi: siklik AMP ve kalsiyumun entegre etkisi. In: Handbook of deneysel farmakoloji (eds. JW Kebabian ve J.Â. Nathanson), 1982, Cilt. 58/11, s. 227-270. Springer, W. Berlin.
Berridge MJ İkinci haberciler olarak inositol trifosfat ve diasilgliserol, Biochem. J., 1984, 220, 345-360.
Berridge MJ İnositol trifosfat ve diasil gliserol ile iyon kanallarının düzenlenmesi , J. exp. Biol., 1986, 124, 323-335.
Berridge MJ, Irvine RF İnositol trifosfat, ! hücresel sinyal transdüksiyonu, Nature, Londra., 1984, 312, 315-321.
Blaustein MP, Ratzlaff RW, Schweitzer ES Presinaptik sinir terminallerinde hücre içi kalsiyumun kontrolü, Fedn Proc. Fed Am. Sox deneyimi Biol., 1980, 39, 2790-2795.
BUss TVP, Douglas RM, Errington ML, Lynch MA Anestezi uygulanmış sıçanların dentat girusundan endojen amino asitlerin uzun vadeli güçlenmesi ve salınması arasındaki korelasyon, J. Physiol., Lond., 1986, 377, 391-408.
Bradley P., Horn G. damgalama: civciv beyninin bazı bölümlerinde kolinerjik reseptör bölgeleri üzerine bir çalışma, Exp. Brain Res., 1981, 41, 121-123.
Bradley P. t Horn G., Bateson PPG Civciv beyninde damgalamanın morfolojik bağıntıları, Neurosci. Lett., 1979, Ek. 3, S.84.
Bradley P., Horn G. imprinting: civciv hyperstriatum ventrale'nin elektron mikroskobik bir çalışması, Exp. Brain Res., 1981, 41, 115-120.
Brown MW, Horn G. Civciv beynindeki nöronal plastisite: görsel deneyimin hiperstriatal nöronlar üzerindeki elektrofizyolojik etkileri, Brain Res., 1979,
142-147.
Cajal SR Histologie du systeme neuroscience de l'homme et des vertebres. 1911 Cilt 2, s. 886-890. Malone, Paris. [Yeniden basım 1955, Histologie du systeme neirux. Instituto Ramón y Cajal, Madrid.]
Carew TJ, Hawkins RD, Abrams TW, Randel ER Aplysia, J. Neurosci., 1984, 4, 1217-1224'te klasik koşullanmaya aracılık eden tanımlanmış sinapslarda Hebb'in varsayımının bir testi .
Apikal dendritlere, Cold Spring Harb'a özel referansla Chang HT Kortikal nöronlar. Semptom nicelik Biol., 1952, 17, 189-202.
Changeux J.-P., Mikoshiba K. Omurgalı beyincik ve nöromüsküler kavşakta sinaps oluşumunu düzenleyen genetik ve "epigenetik" faktörler, Progs. Brain Res., 1978, 48, 43-64.
Cohen MW İn vivo ve kültürde bir amfibi nöromüsküler kavşağının geliştirilmesi, J. exp. Biol., 1980, 89, 43-56.
ColUngridge GL, Kehl SJ, McLennan H. Sıçan hipokampüsünün Schaffer komissural yolundaki sinaptik iletimde uyarıcı amino asitler, J. Physiol. Londra., 1983, 334, 33-46.
Colonnier M. Kedi görsel korteksinin farklı laminalarında farklı hücre tiplerinde sinaptik desenler. Bir elektron mikroskobu çalışması, Brain Res., 1968, 9, 268-287.
Colonnier M., Rossignol S. Serebral korteksin heterojenliği. İçinde: Epilepsilerin temel mekanizmaları (eds. HH Jasper, AA Ward ve A. Pope), 1969, s. 29-40, Küçük, Kahverengi, Boston.
Cragg BG Serebral korteksin motor ve görsel alanlarındaki sinapsların ve nöronların yoğunluğu, J. Anat., 1967, 101, 639-654.
Davies DC, Horn G. Piliç hyperstriatum ventrale'nin varsayılan kolinerjik afferentleri: kombine bir asetilkolinesteraz ve retrograd floresan etiketleme çalışması, Neurosci. Lett., 1983, 38, 103-107.
Downes, C.P. Merkezi sinir sisteminde alıcı tarafından uyarılan inositol fosfolipid metabolizması, Celi Calcium, 1982, 3, 413-428.
Eccles JC Sinapsların fizyolojisi. Springer, W. Berlin, 1964.
Fagg G-E-, Matus A. N-metil aspartat ve quisqualate tip L-glutamat reseptörünün beyin postsinaptik yoğunlukları ile seçici ilişkisi, Proc. nat. Acad. Sci., ABD, G984, 81, 6876-6880.
Fambrough DM Biyosentezi ve nikotinik asetilkolin reseptörlerinin devri. İçinde: İlaç reseptörleri ve efektörleri (ed. NJM Birdsall), 1981, s. 155-
Macmillan, Londra.
Fifkovâ E., van Harreveld E. Entorhinal bölgenin uyarılmasının ardından dentat granüler hücrelerin dendritik dikenlerinde uzun süreli morfolojik değişiklikler, J. Neurocytol., 1977, 6, 211-230.
Garey L. Powell TPS Kedi ve maymunda lateral genikülo-kortikal yolun sonlandırılması üzerine deneysel bir çalışma, Proc. R. Soc., 1971, B179, 41-63.
Goy RW, McEwen BS Beynin cinsel farklılaşması. MIT Press, Cambridge, Mass, 1980.
Grab DJ, Carlin RK, Siekevitz P. Postsinaptik yoğunlukta kalmodulin'in varlığı ve işlevleri, Ann. NY Acad. Bilim 1980, 356, 55-72.
Grab DJ, Carlin RK, Siekevitz P. Kalmodulin'in postsinaptik yoğunluklarda işlevi. II. Kalmodulin ile aktive olabilen bir protein kinaz aktivitesinin varlığı, J. Celi Biol., 1981, 89, 440-448.
Gri EG Serebral korteksin akso-somatik ve akso-dendritik sinapsları: elektron mikroskobik bir çalışma, J. Anat., 1959, 93, 420-433.
Gray EG, Guillery RW Dendritik omurga aparatı üzerine bir not, J. Anat., 1963, 97, 389-392.
Green JD, Arduini A. Uyarılmada hipokampal elektriksel aktivite, J. Neurophysiol., 1954, 17, 533-557.
Griffith JS Belleğin doğasına ilişkin bir teori, Nature, Londra., 1966, 211, 1160-1163.
Griffith JS Beynin bir görünümü. Clarendon Press, Oxford, 1967.
Griffith JS Bellek ve hücresel kontrol süreçleri. İçinde: Kantitatif Metabolizma Biyolojisi (ed. A. Locker), 1968, s. 234-244, Springer, W. Berlin.
Hartzell HC Mechanisms, Nature, Londra., 1981, 291, 539-544.
Hawkins RD, Abrams TW, Carew TJ, Kandel ER Aplysia'da klasik şartlandırmanın hücresel bir mekanizması : presinaptik kolaylaştırmanın aktiviteye bağlı amplifikasyonu, Science, 1983, 219, 400-405.
Hebb DO Davranış organizasyonu, 1949, John Wiley, New York.
Heidman T., Changeaux J.-P. Synapse chimique'in sinaptik sonrası niveau niveau regülasyon etkili bir molekül modeli, C. r. Acad. Sci., Paris, 1982, 295, 665-670.
Horti G. Algının bazı sinirsel bağıntıları. İçinde: Biyolojide bakış açıları (eds. JD Carthy ve CL Duddington), 1962, Cilt. 1, s. 242-285. Butterworth, Londra.
Horn G., McCabe BJ Damgalama: civciv beyninde N-metil-D-aspartat (NMDA) reseptörlerinde lokalize değişiklikler, J. Physiol., Londra., 1987 (baskıda).
Horn G., Bradley P., McCabe BJ Öğrenme ile ilişkili sinapsların yapısındaki değişiklikler, J. Neurosci., 1985, 5, 3161-3168.
Horning JP t Garey LJ Kedi görsel korteksine talamik projeksiyon: Golgi emprenyesi veya retrograd yaban turpu peroksidaz nakli ile tanımlanan nöronların ultra yapısı, Neuroscience, 1981, 6, 1053-1068.
Irvine RF Memeli hücrelerinde serbest araşidonik asit seviyesi nasıl kontrol edilir? biyokimya J., 1982, 204, 3-16.
Jack JJB t Noble D., Tsien RW Uyarılabilir hücrelerde elektrik akımı akışı. Clarendon Press, Oxford, 1975.
James W. Psikolojinin ilkeleri, Henry Holt, New York [Yeniden basıldı (1950) Dover Publications, New York], 1890.
Kehoe J. t Marty A. Belirli yavaş sinaptik yanıtlar: bunların özellikleri ve altında yatan olası mekanizmalar, A. Rev. biyografiler. Bioengng., 1960, 9, 437-465.
Kelly P, T., McGuinness TL, Greengard P. Başlıca postsi naptik yoğunluk proteininin bir Ca2 + /kalmodulin bağımlı protein kinazın bir bileşeni olduğuna dair kanıt, Proc, natn. Acad. bilim ABD, 1984, 81, 945-949.
Kennedy MB, Bennett MK, Erondu NE "Major postsinaptik yoğunluk proteininin" kalmodulin bağımlı protein kinaz Proc'un bir alt birimi olduğuna dair biyokimyasal ve immünokimyasal kanıt. nat. Acad. bilim ABD, 1983, 80, 7357-7361.
Koch C., Poggio T. Dendritik dikenlerin elektriksel özelliklerinin teorik bir analizi, Proc. R. Soc 1983, B218, 455-477.
Kohonen T. İlişkisel bellek: sistem-kuramsal bir yaklaşım, 1984, Springer, W. Berlin.
140
Konorski L Koşullu refleksler ve nöron organizasyonu, 1948, Cambridge University Press, Londra.
Kosower EM Öğrenme ve hafıza için nioleküler bir temel, Proc. nat. Acad. Sci., ABD, 1972, 69, 3292-3296.
Kretsinger RH (ed.). Kalsiyum tarafından seçici sinyalleşme mekanizmaları, Neurosci. Res. Program Bülteni, 1981, 19, 213-328.
Kupfennann L, Carew TJ, Randel ER Aplysia, J. Neurophysiol., 1974, 37, 996-1019'da solungaç ve sifon hareketlerini kontrol eden yerel, refleks ve merkezi komutlar .
Longuet-Higgins HC Holografik zamansal hatırlama modeli, Nature, Londra., 1968, 217, 104.
LUnds R., Sugimori M. Memeli serebellar dilimlerinde in vitro Purkinje hücre dendritlerinin elektrofizyolojik özellikleri. J. Physiol., Londra, 1980, 305, 197-213.
Llinâs R., Yarom U. Memeli alt zeytin nöronlarının in vitro elektro-tepkiselliğini oluşturan iyonik iletkenliklerin özellikleri ve dağılımı, J. Physiol., Lond., 1981, 315, 569-584.
Lynch M, A., Errington ML, Bliss TVP Dentat girusta uzun vadeli sinaptik iletimin güçlendirilmesi: reseptör bağlanmasında artış olmaksızın [ I4C ] glutamat salınımının artması, Neurosci. Lett., 1985, 62, 123-129.
McCabe BJ, Horn G., Bateson PPG Civciv ön beyninin sınırlı lezyonlarının damgalama sırasında evlat tercihlerinin edinilmesi üzerindeki etkileri, Brain Res., 1981, 205, 29-37.
MacDermott AB, Mayer ML, Westbrook GL, Smith JS, Barker JL NMDA-reseptör aktivasyonu, kültürlenmiş spinal nöronlarda sitoplazmik kalsiyum konsantrasyonunu arttırır, Nature, Lond., 1986, 321, 519-522.
McGraw CF, Somlyo A. V. Blaustein MP Presinaptik sinir terminallerinde kalsiyum araştırması, Fedn Proc. Fed Am. Sox deneyimi Biol., 1980, 39, 2796-2801.
Matus A. L, Walters B. B. LJ Memeli beyninden izole edilen sinaptik bağlantı kafesinin yapısı, J. Neurocytol., 1975, 4, 369-375.
Mayhew TM Kantitatif mikroskobik anatomi için temel stereolojik ilişkiler — basit bir sistematik yaklaşım, J. Anat., 1979, 129, 95-105.
Miller JP, Jacobs GA Nöronal yapı ve fonksiyon arasındaki ilişkiler. Sinir sistemindeki entegrasyon mekanizmaları, J. exp. Biol., 1984, 112, 129-145.
Milner P. M. Hücre düzeneği: Mark II, Physiol. Rev., 1957, 64, 242-252.
Neering IR t McBurney RN Memeli nöronlarında mikrozomal Ca depolamasının rolü? Nature, Londra., 1984, 309, 158-160.
Nishizuka W. Protein kinaz C'nin hücre yüzeyi sinyal transdüksiyonu ve tümör teşvikindeki rolü, Nature, Londra., 1984, 308, 603-698.
Payne JKt Horn G. Civciv beyninin iki bölgesindeki "spontane" nöronal aktivite üzerinde damgalayıcı bir uyarana maruz kalmanın farklı etkileri, Brain Res., 1982, 232, 191-193.
Payne JK, Horn G. Civciv beynindeki spontan dürtü aktivitesi üzerinde damgalayıcı bir uyarana maruz kalmanın uzun vadeli sonuçları , Behav. Brain Res., 1984, 13, 155-162.
Peters A., Kaiserman-Abramof IR Sıçan-serebral korteksin küçük piramidal nöronu. Perikaryon, dendritler ve dikenler, Am. J. Anat., 1970, 127, 321-356.
Peters A. t Palay SL, Webster H. de F. Sinir sisteminin ince yapısı: nöronlar ve destekleyici hücreler, 1976, Saunders, Philadelphia.
Pribram KH Hatırlamanın bazı boyutları: nörofizyolojik bir bellek modeline doğru adımlar. İçinde: Makromoleküller ve davranış (ed. J. Gaito), 1966, s. 165-187, Appleton-Century-Crofts, New York.
Rall W. Dendritik dikenler, sinaptik güç ve nöronal plastisite. İçinde: Nöronal aktivitedeki değişikliklere hizmet eden hücresel mekanizmalar (eds. CD Woody, K. A. Brown, TJ Crow ve JD Knispel), 1974, s. 13-21. Beyin Bilgi Servisi, Kaliforniya Üniversitesi, Los Angeles.
Rall W. Dendritik dikenler ve sinaptik güç. İçinde: Nörofizyoloji Çalışmaları (ed. R. Porter), 1978, s. 203-209. Cambridge University Press, Cambridge.
Rall W., Rinzel J. Şube giriş direnci ve bir dendritik nöron modelinin bir dalına giriş için sürekli zayıflama, Biophys. J., 1973, 13, 648-688.
Rastnussen H. Kalsiyum ve cAMP uyaran - tepki eşleşmesi, Ann, NY Acad. Sci., 1980, 356, 246-353.
Rastnussen FL, Barrett PQ Kalsiyum haberci sistemi: entegre bir görünüm, Physiol. Rev. 1984, 64, 938-984.
Shepherd GM Beynin sinaptik organizasyonu (2. baskı). 1979, Oxford University Press, New York.
Sholl DA Serebral korteksin organizasyonu. 1956 Methuen, Londra.
Smilowitz H., Hajian RA, Dwyer J., Feinstein MB Kalsiyum ve kalmodulin tarafından asetilkolin reseptörü fosforilasyonunun düzenlenmesi, Proc. nat. Acad. Sci., ABD, 1981, 78, 4708-4712.
Stent GS Hebb'in öğrenme varsayımı için fizyolojik bir mekanizma, Proc. nat. Acad. Sci., ABD, 1973, 70, 997-1001.
Stewart MG, Rose SPR, King TS, Gabbott PLA, Bourne R. Pasif kaçınma eğitiminin ardından civciv medial hyperstriatum ventrale'deki sinapsların hemisferik asimetrisi: stereolojik bir araştırma, Dev. Brain Res., 1984, 12, 261-269.
Tanzi E. I fatti e le le induzioni nell'odierna istologia del systema nervosa, Riv. sp. Freniat. Med. bacak Uzaylı. ment., 1893, 19, 419-472.
Valverde F. Farede görsel korteksin apikal dendritik dikenleri ve ışık yoksunluğu, Exp. Brain Res., 1967, 3, 337-352.
Valverde F. Farenin görsel korteksindeki karanlık yetiştirmeden kurtulma derecesi, Brain Res., 1971, 33, 1-11.
Walter's ET, Bryne JH Tek duyu nöronlarının ilişkisel koşullanması, öğrenme için hücresel bir mekanizma önerir, Science, 1983, 219, 405-408.
Watanabe M., Ito H., Japon bıldırcınlarının (Coturnix coturnix-japonica) Wulst'unda Masat H. Cytoarchitecture ve görsel alıcı nöronlar , J. comp. Neurol., 1983, 213, 188-198.
Weibel ER, Bolender RP Elektron mikroskobik morfometri için stereolojik teknikler. İçinde: Elektron mikroskobunun ilkeleri ve teknikleri (ed. MA Hayat), 1973, Cilt. 3, Biyolojik uygulamalar, s. 237-296. Van Nostrand Reinhold, Londra.
Westrum LE, Jones DH, Gray EG, Barron J. Mikrotübüller, dendritik dikenler ve spink aparatlar, Celi Tissue Res., 1980, 208, 171-181.
Yamamoto KR, Alberts B. M. Steroid reseptörleri: ökaryotik transkripsiyonun modülasyonu için elementler, A. Rev. Biochem., 1976, 45, 721-746.
pmvg bir "BELLEK YAPISI" MIDIR!
Çalışma sırasında, damgalama mekanizmaları arayışı yavaş yavaş küçük bir alanla - PMVP ile sınırlandırıldı. Bu alana olan ilgi arttıkça diğer alanlara olan ilgi azaldı . Ancak, SMVG boşlukta çalışmaz . Bu alanın damgalamanın gelişimindeki rolünü anlamak için, organizasyonunu, ona giren bilginin doğasını ve beynin diğer bölümleri üzerindeki etkilerini ayrıntılı olarak bilmek gerekir. Bu bölümün amacı, bu konular hakkında az bilinenleri gözden geçirmektir.
İletişim SMVG'si
Merkezi sinir sistemindeki yolları belirlemenin birçok yöntemi vardır. Yöntemlerden biri, belirli bir bölgeye zarar vermek ve bunun sonucunda sinir liflerinin ve uçlarının dejenerasyonunun izini sürmektir. Diğer bir yöntem ise nöronları zayıf bir elektrik akımıyla uyarmak ve beynin diğer bölümlerindeki kısa gecikmeli uyarılmış tepkileri ortaya çıkarmaktır . Ayrıca , nöronların süreçleri boyunca maddelerin iki yönlü hareketinin kullanımına dayanan, nispeten yakın zamanda geliştirilmiş yöntemlerimiz var - akson terminallerine (anterograd) veya terminallerden perikaryaya (retrograd). Tavuklarda PMCH ilişkilerini analiz etmek için bu yöntemleri kullandık (Bradley ve diğerleri, 1985).
yaban turpu peroksidazı (HRP) içeren bir solüsyonu akson sonlanma bölgesine enjekte eden ve bu enzimin bu aksonların ait olduğu hücre gövdelerinde biriktiğini bulan Christensson ve Olsson (Kristensson ve Olsson, 1971) tarafından önerildi. . Christensson ve Olsson'un çalışması periferik sinir sistemi üzerinde gerçekleştirildi, ancak yöntem kısa süre sonra merkezi sinir sistemi çalışmasına uygulandı (LaVail, LaVail, 1972). Anestezi uygulanmış bir hayvanın beynine az miktarda HRP solüsyonu enjekte edilir. Beynin enjeksiyonu ile fiksasyonu arasındaki aralık , 260 enziminin taşınması gereken mesafeye ve taşıma hızına bağlıdır. Fiksasyondan sonra doku dondurulur, kesilir ve uygun substratlarla inkübe edilir. Kesitler daha sonra daha fazla boyanabilir. HRP reaksiyon ürünü, hücre gövdesinde koyu renkli granüller şeklinde görülür . HRP, var olan tek retrograd belirteç değildir. Nükleer sarı ve güçlü mavi floresan boyalar gibi diğer belirteçler, perikaryondaki farklı yapıları etiketleme avantajına sahiptir. Özellikle güçlü mavi, hücrenin sitoplazmasını boyarken, nükleer sarı hücrenin çekirdeğinde yoğunlaşmıştır (Bentivolgio ve diğerleri, 1980). Bu belirteçlerden birini veya her ikisini içeren hücre gövdeleri, ultraviyole ışık altında floresandır.
Bu yöntemlerin kullanımında bir takım sınırlamalar vardır . Belirteç, enjeksiyon alanının dışına yayılabilir ve sonuç olarak, araştırmacının ilgilendiği bölgeye bağlantı göndermeyen perikaryonlara taşınabilir . Enjeksiyon bölgesinden geçen lifler ( "geçiş" lifleri) , işaretleyiciyi sokmak için kullanılan kılcal damar tarafından zarar görebilir . Aksonlarının enjeksiyon bölgesinde terminalleri olmamasına rağmen, bu liflerin ait olduğu hücre gövdeleri lekeli olabilir. Retrograd floresan boyalar perikaryadan yayılabilir ve komşu hücreleri boyayabilir. Araştırmacının ilgi alanına bitişik alanlarda markörün kontrol enjeksiyonları, markör enjeksiyonu için farklı anatomik yaklaşımlar, enjeksiyon süresi ile fiksasyon arasındaki farklı aralıklar ve farklı markörler kullanılarak bu zorluklar aşılabilir .
anterograd belirteçler kullanılarak incelenebilir . Nöronlar , süreçler boyunca perikaryondan taşınan proteinleri sentezler. Amino asitler hücre gövdelerinin yakınına getirilirse, perikarya tarafından yakalanır ve sitoplazmada hareket eden proteinlere dahil edilir. Kullanılan anterograd yöntemlerle, amino asitlerin proteinlere dahil edilmesi, enjeksiyon bölgesinin yakınında bulunan aksonlarda veya sinapslarda değil, yalnızca hücre gövdeleri bölgesinde gerçekleşir. Yöntem aşağıdaki aşamaları içerir: 1) radyoaktif olarak etiketlenmiş bir amino asidin beynin herhangi bir bölümüne enjeksiyonu; 2) işaretli habercinin dahil edilmesi ve bu bölgenin hücrelerinden çıkan aksonların terminallerine taşınması için gereken çeşitli sürelerde bir aralık ; 3) beyin bölümlerinin tespiti ve hazırlanması ; 4) gümüş tanelerini tespit etmek için radyootografik yöntemin kullanılması (s. 87). Granüllerin konumu daha sonra beynin histolojik bölümleriyle ilişkilendirilir.
140
ortaya çıkan sorunlardan bazıları, retrograd yöntemlerle ortaya çıkanlara benzer; bunlar, örneğin , etiketin enjeksiyon bölgesinden difüzyonunun yanı sıra, birleşik amino asitlerin hasarlı aksonlar tarafından alınması ve taşınmasıdır. Ek olarak, bu durumda , akson terminaline ulaşan radyoaktif öncünün sinaps yoluyla postsinaptik hücrenin perikaryonuna girmesi ve onu işaretlemesi tehlikesi vardır . Bunlar ve diğer bazı güçlükler , en azından kısmen, geriye dönük işaretleyicilerin kullanımıyla bağlantılı olarak belirtilen bazı kontrol prosedürleri kullanılarak aşılabilir.
Bu alana afferent liflerin kaynaklarını ve yansıtıldığı yapıları belirlemek için ışıkta tutulan bir günlük civcivlerin PMVG'sine retrograd veya anterograd belirteçler yerleştirildi (Bradley ve diğerleri, 1985). Bu çalışma gerekliydi çünkü genel olarak ventral hiperstriatum ve özel olarak PMSG arasındaki bağlantılar hakkında çok az şey biliniyor.
Duyusal sistemler ve beyin sapı ile bağlantılar
Görsel sinyallerin SMVG'ye ulaşabileceği dört yol vardır. İlki Wulst'u içerir (Şek.
L). Adamo (1967) , tavuklarda Wulst'a zarar verdi ve medial ventral hiperstriatumda dejenere terminaller buldu. Adamo'ya göre, bu projeksiyon alıcı bölgenin SMVP'yi yakalayacak kadar geriye uzanıp uzanmadığını belirlemek mümkün değil . Böyle bir projeksiyonun varlığına ilişkin veriler , PMVP'ye eklenen retrograd bir işaretleyici kullanılarak elde edildi . Etiketli hücre gövdeleri , Wulst'un tüm katmanlarında bulundu : ek hiperstriatumda (hyperstriatum accessorium), orta hiperstriatumda (hyperstriatum intercalatus) ve dorsal hiperstriatumda (hyperstriatum dorsale). Ek hiperstriatuma retrograd bir işaretleyici eklendiğinde, PMVG'de etiketli hücre gövdeleri tespit edildi. Bu nedenle, PMSH karşılıklı olarak ek bir hiperstriatum ile ilişkilidir. Wulst karmaşık bir görsel projeksiyon alanı olduğundan (Şekil A.5b ) ve görsel uyaranlara yanıt veren nöronlar içerdiğinden (Bölüm A.3.1), Wulst'tan SMVG'ye olan projeksiyonların görsel sinyaller taşıması oldukça olasıdır .
Görsel bilginin PMVP'ye iletilebildiği ikinci yol neostriatumdur. Parker ve Delius (Parker ve Delius, 1972), ışık parlamalarına yanıt olarak , hücre deşarjlarının ve yüksek genlikli indüklenenlerin olduğunu bulmuşlardır.
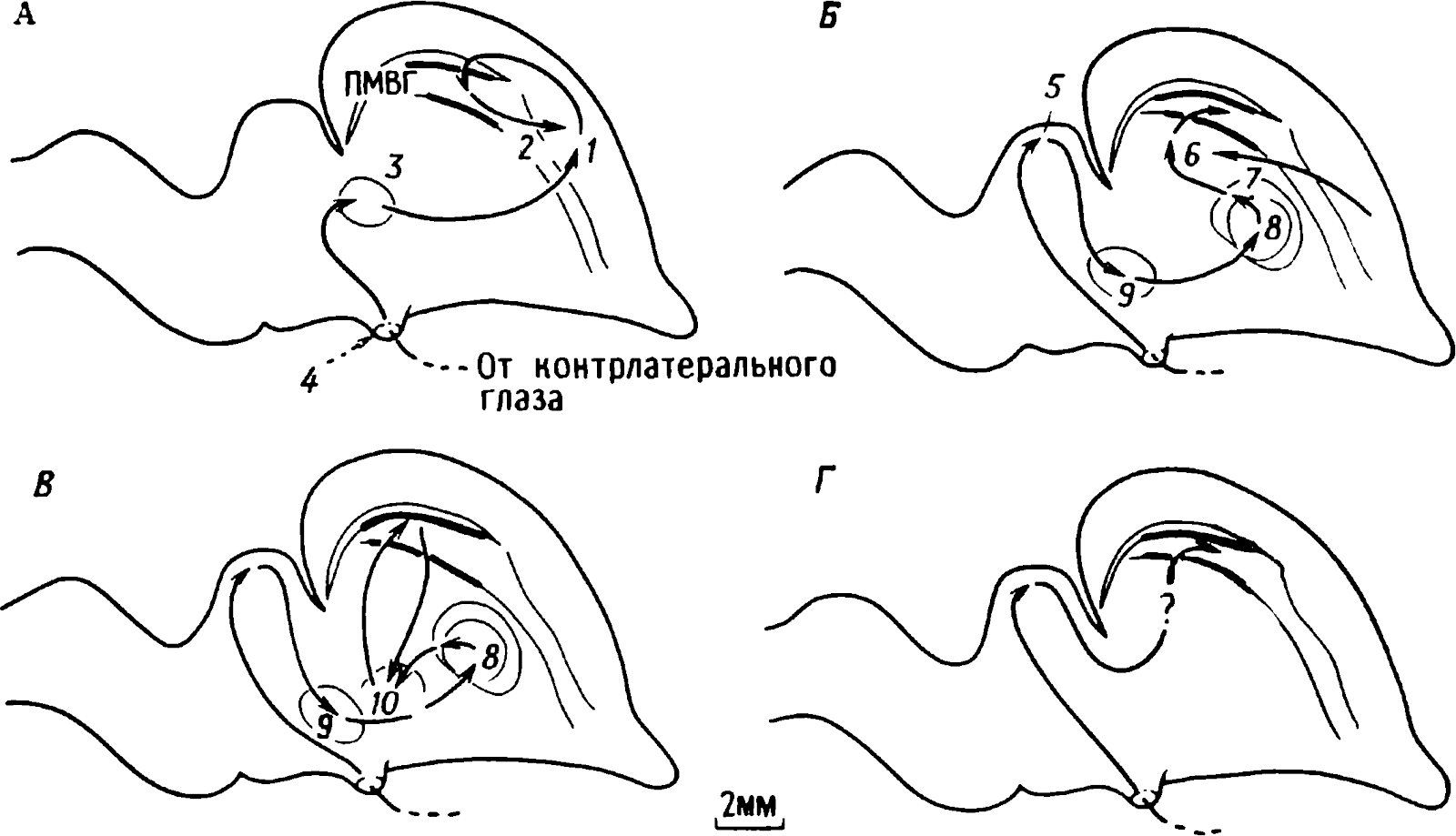
Resim: 11.1. PMVP'ye olası görsel sinyal yolları. A. Vücutları görsel Wulst'un (1) her üç hücre katmanında bulunan nöronlar, PMWG'ye yansıtılır. PMSH ve aksesuar hiperstriatum arasında karşılıklı bağlantılar vardır. Wulst dürbün girişi almasına rağmen, yalnızca kontralateral gözden gelen yollar gösterilir. B. SMVG'ye afferent lifler gönderen neostriatum (6) bölgesi, ek hiperstriatum ve ektostriyal kuşaktan (7) çıkıntılar alır. Bu son alan görsel sistemin tektofugal bileşeninin bir parçasıdır (Bölüm A.3.1). C. Periectostriatal kuşak ayrıca orta arkistriatumun (10) dorsal kısmına da çıkıntı yapar . Bu yapı karşılıklı olarak PMVP ile ilişkilidir. D. Optik tavandaki (5) hücre gövdeleri , PMWG'ye eklenen geriye dönük işaretler içerir. Bu yolla ilgili bazı belirsizlikler var (metne bakın). Bu bölümde ve bu bölümdeki diğer şekillerde gösterilen beyin dilimleri şematiktir; ince bir histolojik kesit üzerinde birlikte görülmeyen kesitler gösterebilirler. Diğer tanımlamalar: 2 - ventral hiperstriatum (PMWG'yi oluşturan kısmı kalın çizgilerle gösterilmiştir; 3 - talamusun ana görsel çekirdeği; 4 - optik yol; 8 - ektostriatum; 9 - yuvarlak çekirdek. Ölçek bu ve diğer şekillerde beynin konturunu ifade eder.
potansiyeller. Boz kazında (Anser anser) bu alanda benzer tepkiler kaydedilmiştir (Felix ve diğerleri, 1983). Görsel sinyallerin neostriatuma girmesinin en az iki yolu vardır (Şekil 11.1, B ). Ek bir hiperstriatum, tavuklarda (Adamo, 1967) ve Amerikan mağara baykuşlarında (Speotyto cunicularia) (Karten ve diğerleri, 1973) neostriatum üzerine projekte olur. Ritchie ve Cohen (1977) , periektostriat kuşağının güvercinlerde ara neostriatumun yanal kısmına bağlantılar gönderdiğini bulmuşlardır. Neostriatumun orta ve kaudal kısmındaki sinir hücreleri, SMVG'ye bağlantılar gönderir (Bradley ve diğerleri, 1985).
18. _
Görsel sinyalleri SMVG'ye iletebilen üçüncü multisinaptik yol, Şekil 1'de gösterilmiştir. 11.1, Ritchie ve Cohen ( 1977) , periectostriatal kuşağın ayrıca ara arşistriatumun dorsal kısmına da çıkıntı yaptığını bulmuşlardır. Bu alan ayrıca PMVP ile karşılıklı bağlantılara sahiptir.
PMVP'ye giden daha doğrudan bir görsel yol, görsel çatının derin katmanında bulunan hücrelerden geliyor gibi görünmektedir (Şekil 1).
G). HRP, PMVP'ye eklendiğinde, optik çatıdaki hücreler etiketlendi. Bu sonuç söz konusu yapılar arasında talamusta bir ara sinaps olmaksızın bağlantı olduğunu göstermektedir. İşitsel (Delius ve diğerleri, 1979), dokunsal (Wallenberg, 1903) ve koku alma (Rieke ve Winzel, 1978) sinyallerinin talamusta geçiş yapmadan ön beyne ulaştığına dair ikna edici kanıtlarımız olmasına rağmen , benzer bir görsel yol daha önce görülmemişti. rapor edilmiştir. hiçbir omurgalıda tarif edilmemiştir . Bu nedenle deneyler, retrograd belirteçler olarak floresan boyalar kullanılarak tekrarlandı. Bu belirteçler ayrıca SMVG'ye sokulduğunda optik çatının hücrelerine de taşınmıştır. Etki oldukça belirgindi. PMCH'ye bitişik yapılara sokulan retrograd belirteçler , optik çatıda tespit edilmedi. Bu yol hakkında ek bilgi elde etme girişiminde optik çatıya anterograd bir boya enjekte ettik . Etiket, optik çatı nöronlarının bilinen hedeflerinde bulundu, ancak etiketli terminaller, PMVP'de saptanmadı; Dolayısıyla geriye dönük ve ileriye dönük yöntemlerle elde edilen sonuçlar arasında bir tutarsızlık vardı . Bu tutarsızlık, yalnızca optik çatıdan SMVG'ye olan projeksiyonların dağınık olması ve geniş bir terminal akson dallanması olan az sayıda nöron tarafından oluşturulması durumunda açıklanabilir. Anterograd yöntemini kullanarak, terminallerin bu tür bir dağılımını tespit etmek kolay değildir .
Şek. 11.1 görsel sinyalleri PMSH'ye iletmezler, farklı özelliklere sahip olmalıdırlar. Ne olabilirler, sadece tahmin edilebilir. Tipik olarak, duyusal sinyaller sinapslardan geçerken değiştirilir ve yolları boyunca ne kadar çok sinaps olursa, modifikasyon o kadar büyük olur. Örneğin, kedinin retinasındaki gangliyon hücreleri, dağınık ışık parlamalarına yanıt verir. Bu hücrelerin aksonları , hücreleri retina ganglion hücrelerinden daha az yaygın flaşlara yanıt veren lateral genikulat gövdede sinapslar oluşturur . Yanal genikülat vücut, görsel korteks üzerine çıkıntı yapar. Görsel korteksin hücreleri parlak ışık parlamalarına çok zayıf tepki verir; onlar için
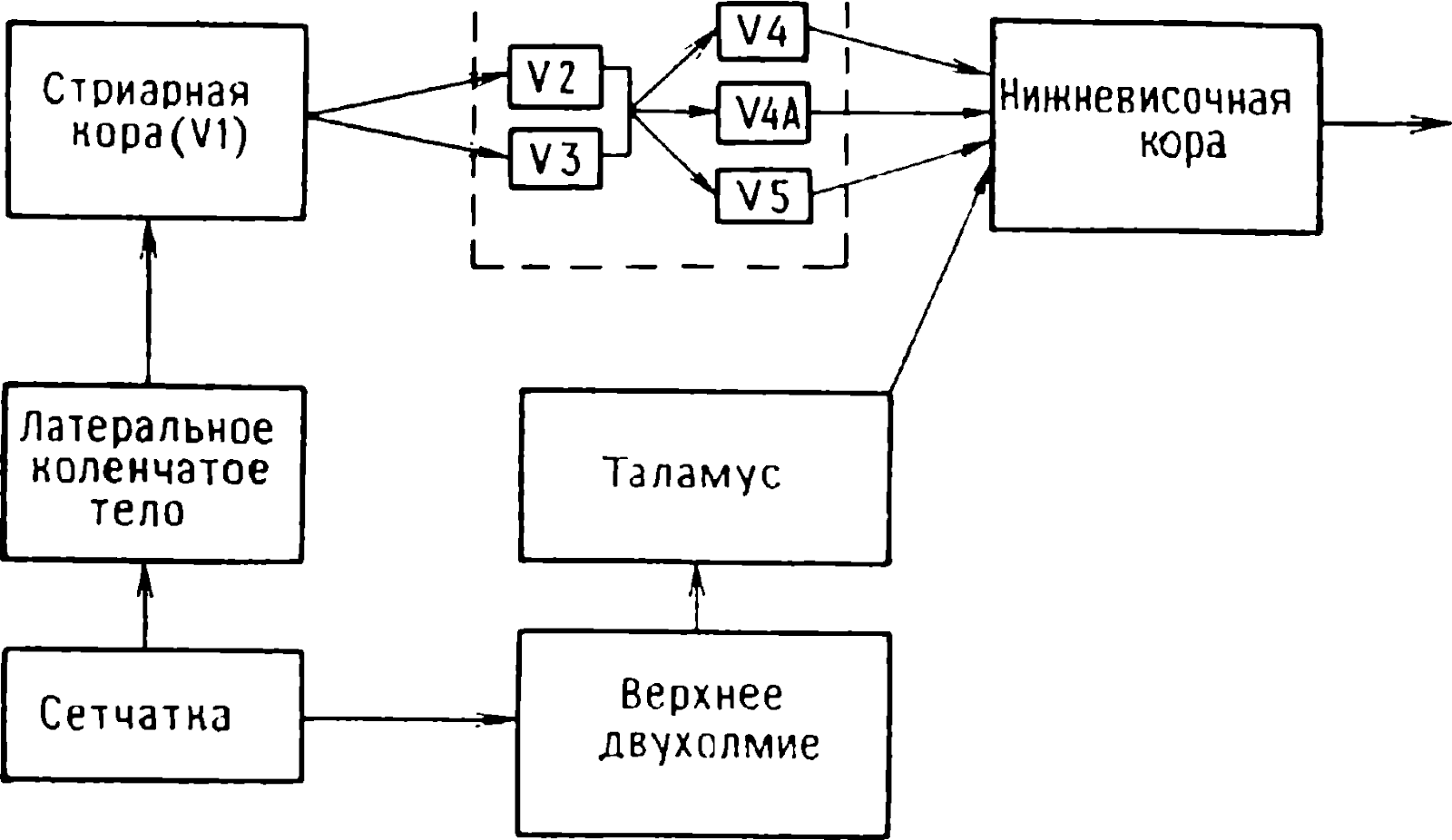
belirli özelliklere sahip uyaranlara ihtiyaç vardır. Örneğin , bu tür nöronlar, alıcı alanları içindeki bir çizgiye yanıt verebilir . Yanıtın büyüklüğü, kritik olarak hattın genişliğine ve yönüne, uzunluğuna, yönüne ve hareket hızına bağlı olabilir (bkz . Hubel tarafından yapılan inceleme, 1982). Bir kedinin görsel sistemi ile bir benzetme yaparak, Şekil 1'de gösterilen multisinaptik yollarda sinyal dönüşümünün olması beklenebilir . 11.1,4 - B, şekil 2'de gösterilen oligosinaptik yoldan daha büyük olmalıdır. 11.1, G.
Aynı yapı içinde multisinaptik ve oligosinaptik yolakların birleşmesi yaygın olarak kabul edilen bir ilkedir ve bu yapıya gelen duyusal bilginin işlenmesi için önemli çıkarımlara sahip olabilir. PMIG'nin bu bölgenin bir homologu olduğuna hiçbir şekilde inanmasak da , bu tür sonuçların varlığına ilişkin veriler, al yanaklı maymunun inferotemporal korteksi üzerinde yapılan deneylerde elde edildi . Alt temporal korteks, bazı açılardan SMVG'ye giden yollara benzeyen geniş bir görsel girdiler grubuna sahiptir (Şekil 11.2). Görsel korteksten geçen yol, alt-temporal kortekse ulaşmadan önce en az beş sinaps (bunlardan en az ikisi retinadadır) içerir. Bu sayı, Şekil 1'de gösterilen SMVG'ye giden yollarda bulunan sayı ile yaklaşık olarak çakışmaktadır. 11.1,4- B. Alt temporal kortekse giden daha doğrudan yol, optik çatıdan geçer (superior kollikulus, Şekil 11.2). Bu alandaki nöronlar doğrudan alt temporal kortekse yansımaz: yol , talamusta en az bir sinaps içerir. Şekil l'de gösterilen yola benzer. 11.1, D, talamik sinaps içermemesine rağmen. Rhesus maymununda , talamusun optik kollikulus ve pulvinarından geçen multisinaptik yol ve oligosinaptik yol , alt temporal korteksteki nöronların alıcı alanlarının özelliklerini farklı şekillerde etkiler . Yastık yok edildiğinde, aşağı temporal kortekste pratik olarak tek bir nöron bile ayrı alıcı alanları tutmaz; çizgili korteksin yıkımı, alıcı alanların uyarılmasına verilen yanıtların özelliklerini değiştirir (Gross ve diğerleri, 1974). Bu temelde, alıcı alanların uzamsal kapsamının yastıktan oligosinaptik yollarla belirlendiği, nöron yanıtlarının özelliklerinin ve bunların uyaran özelliklerine bağımlılığının kortiko-kortikal multisinaptik girdi tarafından belirlendiği sonucuna varıldı.
İnferotemporal rhesus korteks, her iki yapının da çoklu ve oligosinaptik yollar alması bakımından tavukların PMIG'sine benzese de, bu benzetme abartılmamalıdır. Alt temporal korteksin iki taraflı çıkarılması, görsel uyaranların farklılaşmasında önemli bir bozulmaya yol açar. Bu bozulma, boyut, şekil ve renk bakımından değişen çok çeşitli uyaranlarla ortaya çıkar ve çok çeşitli öğrenme yöntemlerinde bulunur (Gross, 1973). Bunun aksine, görsel olarak yönlendirilen birçok davranış biçimi, PMSG'nin zarar görmesinden etkilenmez (Bölüm 6 ve Kısım 8.4.1).
Çoklu ve oligosinaptik görsel yolların SMVG'ye etkileşiminin benzer bir analizi henüz yapılmadı. Gerçekten de, ventral hiperstriatumun daha geniş alanındaki nöronların görsel tepkilerinin özellikleri bile çok az çalışılmıştır. Morenkov ve Hun (1977) , gri karganın (Corvus corone) ventral hiperstriatumunun medial kısmındaki nöronların aktivitesini kaydetti. Görünüşe göre PMCH'ye anatomik olarak karşılık gelen bölgedeki nöronlar, 30-50° çapa ulaşan nispeten geniş alıcı alanlara sahipti. Bu nöronlar, belirli bir yönde hareket eden uyaranlara tercihli olarak yanıt verdiler, yani hareket yönüne doğru seçiciydiler. Guselnikov ve diğerleri (1976) , yetişkin güvercinlerde ventral hiperstriatum nöronlarının aktivitesini kaydetmiştir. Bu hücrelerin tepkileri ayrıntılı olarak açıklanmamıştır , ancak resimlerden ventral hiperstriatumun ön kısmının, uyaran hareketinin yönüne göre seçici olan yerel alıcı alanlara sahip hücreler içerdiği açıktır. Mikroelektrotlar , anatomik olarak PMSH'ye karşılık gelen alana yerleştirilmedi. Buna karşılık, Wilson ( 1980), 5 haftalık civcivlerde aksesuar hiperstriatumda görsel uyaranlar tarafından indüklenen aktiviteyi tespit edemedi . Uyaran olarak, hareketsiz nesnelerin yanı sıra hareketli ışık noktaları, şeritler ve dikdörtgenler kullanıldı. Wilson tarafından incelenen ventral hiperstriatum alanı, aksesuar hiperstriatumdan daha derindir ve bu nedenle PMH'yi içermez. Ancak Milne, Horne ve McCabe (yayınlanmamış gözlemler), karanlıkta tutulan 2 günlük civcivlerde ve 2 haftalık olana kadar görme deneyimi olan civcivlerde PMVP nöronlarının aktivitesini kaydetti. Wilson tarafından kullanılanlara benzer uyaranlar kullanıldığında , az sayıda hücre dağınık ışık flaşlarına zayıf tepki vermesine rağmen görsel yanıt bulunamadı (Brown ve Hogn, 1979).
Bu sonuçların tutarsızlığı net değil, ancak birkaç nedenden dolayı şaşırtıcı değiller ve tutarsızlıkları gerçek olmaktan çok aşikar olabilir. Ayrık görsel uyaranlara nöronal tepkiler bulan iki çalışmada yetişkin güvercinler ve kargalar kullanılmış, evcil tavuklarda ise negatif sonuçlar elde edilmiştir. PMVG'nin görsel uyarıya gösterdiği reaktivitede türler ve yaş farklılıkları olabilir. Ek olarak, Morenkov ve arkadaşları (1976) anestezi uygulanmamış kuşlar üzerinde çalıştılar; tavuklara anestezi uygulandı. Deneylerin tasarımındaki bu farklılık önemli olabilir . Duyusal stimülasyona verilen nöronal tepkilerin , özellikle incelenen alan duyu reseptörlerinden sadece birkaç sinaps uzaktaysa , anestezi uygulanmış hayvanların ana duyusal yollarında tespit edilmesi zor değildir . Buna karşılık, anestezik ajanlar genellikle multisinaptik yollar üzerinde güçlü bir baskılayıcı etkiye sahiptir (Clutton-Brock, 1961; Brazier, 1961; Brown ve Horn, 1977). Bu nedenle, duyu organlarından sinaptik olarak uzak olan bölümlerden gelen impulsları kaydederken anestezikler genellikle kullanılmaz. Bununla birlikte, beynin elektriksel aktivitesini kaydederken bazen anesteziyi bırakmanın gerekli olmasının başka bir nedeni daha vardır . Bu ajanlar, dikkat, öğrenme veya karmaşık ve hatta basit motor eylemler gibi davranışın neredeyse tüm ilginç yönlerini ortadan kaldırır . Bundan anestetiklerin beynin bu işlevleri sağlayan nöral aktiviteyi bütünleştiren kısımlarını etkilediği sonucu çıkar. Bu nedenle , beynin bu işlevlerle ilgili olabilecek bölümlerinin modern elektrofizyolojik çalışmalarının yürütülüyor olması şaşırtıcı değildir.
Resim: 11.3. PMVG'ye giden işitsel yollar. Sekizinci kraniyal sinirin işitsel kısmının aksonları , beyin sapının işitsel çekirdeklerinde son bulur. Aşağıdaki yollar düz ve çapraz bileşenleri içerir. Ön beyindeki birincil projeksiyon işitsel alanı , kaudal neostriatumdaki L alanı ile temsil edilir. Bu alanın ön kısmı LMVH ile örtüşür, ancak L alanı LMVG'nin en kaudal kısmının ötesinde posterior olarak devam eder. L alanı, Lı yukarıda ve L3, L2'nin altında olacak şekilde üç kısma bölünmüştür . L alanı PMVP'ye yansıtılır. 1 - sekizinci kranial sinirin işitsel kısmı ; 2 - medulla oblongata'nın işitsel çekirdeği; 3 - orta beynin dorso-lateral çekirdeği; 4 - oval çekirdek; L - neostriatumdaki L alanı. Kesikli çizgiler beynin kontralateral yapılarını, düz çizgiler ipsilateral olanları gösterir. Daha ayrıntılı açıklamalar için bkz. S.2.
serbest davranışta anestezi uygulanmamış hayvanlar üzerinde (bakınız, örneğin, Wurtz ve Goldberg, 1971; Hoot ve Wiesenfeld, 1974; Mountcastle, 1976; Mora ve diğerleri, 1976; Brown ve Hoot, 1978; Wurtz ve diğerleri, 1980).
PMVG'sinde spesifik alıcı alanlara sahip hücreler bulunmamakla birlikte, bu alandaki nöronların spontan aktivitesinin ve sinapsların yapısının, imprinting gelişimine neden olan görsel bir uyaranın etkisi altında değiştiği vurgulanmalıdır (Bölüm 10).
Ön beynin ana işitsel projeksiyon alanı , neostriatumun kaudal kısmında yer alan L alanıdır (Bölüm A.3.2 ve Şekil A.6). Tavukta L alanı ventral hyperstriatum'un medial kısmına bitişiktir ve ondan lamina hyperstriatica ile ayrılır (Scheich, 1983). L alanı , LMVP ile kısmen örtüşür ve sınırlarının ötesinde posterior olarak devam eder. Topografik ilişkiler, Şek. 11.3. İşitsel projeksiyon alanı, neostriatumun görsel projeksiyon alanına medial olarak uzanır (Parker ve Delius, 1972). Sheikh (Scheich, 1983), L alanını rostro-kaudal yönde üç fonksiyonel bölgeye ayırdı. Muhtemelen PMIG ile örtüşen ön ve orta kısımların kokleadan iki taraflı afferent yollar aldığını gösterdi. Bu organizasyon, bu alandaki nöronların sesin lokalizasyonunda yer aldığını ima eder. Ayrıca türe özgü seslendirmeleri de işleyebilirler (Scheich, 1983; Scheich ve diğerleri, 1979b). Bunun böyle olup olmadığına bakılmaksızın, L alanının ön ve ara kısımlarındaki nöronların karmaşık ses sinyallerini analiz edebildiği deneysel olarak gösterilmiştir. Civcivlerde, L alanından gelen bağlantılar PMVG'ye gider. Retrograd belirteçler PMIG'e eklendiğinde , bitişik neostriatumda, L alanına karşılık gelen alanda etiketli hücre gövdeleri bulundu (Bradley ve diğerleri, 1985). Beç tavuğunda, L alanı ve bitişik ventral hiperstriatum karşılıklı yollarla bağlanır (Bonke ve ark.,
.
Evcil tavuğun ventral hiperstriatumunun medial kısmındaki nöronların basit sesli uyaranlara yanıt vermemesi mümkündür. Scheich ( 1983) , civcivler frekans modülasyonlu tonlarla beslendiğinde radyoaktif 2-deoksiglukoz (Bölüm 4.2) dahil ederek nöronal aktiviteyi değerlendirdi . Bu stimülasyon L alanında güçlü bir etki yarattı , ancak yayınlanan fotoğraflar bunun ventral hiperstriatumdaki metabolizma artışıyla ilişkili olmadığını gösteriyor (Scheich, 1983). Beç tavuğu saf tonlara maruz kaldığında, L alanında da yoğun işaret birikimi gözlendi , ancak ventral hiperstriatumda çok zayıftı (Scheich ve diğerleri, 1979a ). Bununla birlikte, bu kuşlar türe özgü seslere maruz kaldıklarında , içinde 2-deoksiglikoz alımının net bölgeleri belirdi . Bu bölgenin, türe özgü sesler de dahil olmak üzere karmaşık işitsel uyaranlar tarafından güçlü bir şekilde aktive edilmesi özellikle ilgi çekicidir. Gottlieb ( 1971), yaban ördeği yavrularında aşağıdaki tepkinin , doldurulmuş bir ördeğin sunumuna karmaşık seslerin eşlik ettiği durumlarda, hiç sesin olmadığı duruma göre daha kolay ortaya çıktığını gösterdi. Ek olarak, diğer bölümlerde anlatılan tavuklar üzerindeki deneylerin birçoğunda (bkz. örneğin, Hom ve diğerleri, 1979; Davies ve diğerleri, 1985), eğitim süresi boyunca görsel-işitsel kombinasyonlar kullanıldı. Bu prosedürün görsel bir uyarana damgalamanın etkinliğini arttırdığı bulundu. Karmaşık uyaranın artan etkinliğinin açıklamalarından biri , sesin kuşun dikkatini görsel nesneye çekmesidir (Gottlieb ve Klopfer, 1962). Başka bir olasılık, karmaşık bir işitsel uyaranın görsel bir uyaranla kombinasyonunun, PMVP nöronları üzerinde bu uyaranlardan herhangi birinin tek başına olduğundan daha güçlü bir etkiye sahip olmasıdır. Bu sorulara kesin cevaplar , serbest davranıştaki tavuklarda bu alandaki nöronal reaksiyonların özelliklerinin mikroelektrot analizi ile verilmelidir .
hafif bir dokunuşla , tüylerin mekanik olarak yer değiştirmesiyle veya tüylere yönelik hava yaylımlarıyla uyandırılan somatosensoriyel tepkiler , neostriatumun kaudal kısmındaki güvercinlerde kaydedilebilir (Şekil 11.4).
Resim: 11.4. PMVG'ye giden somatosensoriyel yollar. Omurilikten gelen duyu lifleri talamusun (1) dorsal çekirdeğine birkaç yoldan gider. Bir yol hassas ve sfenoid demetlerin çekirdeklerinde bir sinapsa sahiptir (2). Beşinci kraniyal sinir (V), başın ön yüzeyinden ve gagadan beyin sapına sinyaller iletir ve ana duyu çekirdeğinde sonlanır (3). Bu çekirdekte, kısmen kesişen bazal çekirdekte biten quintofrontal yol başlar (4). Delius ve Bennetto ( 1972), bu yolun bir kısmının dorsal talamusta bittiğine inanırlar. Onlara göre bu çekirdek neostriatumun somatosensoriyel bölgesine yollar gönderir (5). Bu bölgenin hücreleri PMVP'ye yansıtılır. Kesikli çizgi ile işaretlenmiş sinirler, yollar ve çekirdekler kontralateral, düz çizgi olanlar ipsilateraldir. Traktları gösteren çizgilerin yanındaki soru işaretleri, elektrofizyolojik çalışmalara dayanarak varlığı varsayılan yolları gösterir. (Delius ve Bennetto, 1972.)
Bu izdüşüm çoğunlukla kontralateraldir (Erulkar, 1955; Delius ve Bennetto, 1972 ). Bu araştırmacılara göre, somatosensori alan işitsel projeksiyon bölgesinin önünde ve arkasında yer alır. Gri kazın kaudal neostriatumunda da somatosensoriyel uyarıma tepkiler kaydedilmiştir (Felix ve diğerleri, 1983). Tavukta bu alan, PMSH'yi içeren ventral hiperstriatum alanına yansıtılır (Bradley ve diğerleri, 1985). Erulkar'ın ( 1955) çalışmasında, somatik uyarılmış yanıtların kaydedildiği bazı noktalar ventral hiperstriatumun medial kısmındaydı.
Koku alma sisteminden ventral hiperstriatuma giden yol , neostriatumdan geçmez (Şekil 11.5). Güvercinlerde, koku ampullerinin hasar görmesi , ipsilateral ventral hiperstriatumda, PMSH'ye karşılık gelen kısmı da dahil olmak üzere terminal dejenerasyona yol açar (Rieke ve Wenzel, 1978). Bu yapıdaki koku alma sinirinin uyarılması iki taraflı yanıtlara neden olabilir (Macadar ve ark. 1980). Toplamda, bu sonuçlar çaprazlanmamış yolun koku ampulünden sinaptik anahtarlama olmadan ventral hiperstriatuma ilerlediğini gösterir. Çapraz yol
Resim: 11 5. Mavi noktadan ventral hiperstriatuma giden koku yolları ve innervasyon. Güvercinlerde, koku ampulünün (1) nöronları , PMSH'ye karşılık gelen alan da dahil olmak üzere ventral hiperstriatum üzerine çıkıntı yapar. PMVG, norepinefrin içeren sinir lifleri içerir; muhtemelen mavi noktada (2) ve bitişik alanlarda bulunan nöronlara aittirler (ayrıca bkz. Şekil A.7).
bu iki yapı arasında en az bir sinaps vardır.
Bu çalışmalardan elde edilen veriler, evcil tavuklarda PMVP'nin veya diğer kuş türlerinde buna karşılık gelen bölgenin tüm duyusal modalitelerden sinyaller aldığını göstermektedir . Tat uyaranlarını ön beyne taşıyan yollar hakkında çok az şey bilinmesine rağmen, tat hassasiyeti tek istisnadır (Gentle, 1975); aynısı vestibüler sistemin merkezi bağlantıları için de geçerlidir. Geri kalan duyusal yolların PMCH'ye olan projeksiyonları hakkındaki veriler esas olarak anatomik çalışmalardan elde edilir, bu nedenle bu sinyallerin etkisinin kesin doğası hala elektrofizyolojik yöntemler kullanılarak açıklanmalıdır.
Duyusal sistemlerden gelen bağlantılara ek olarak, PMH şüphesiz beyin sapının merkezi yapılarından yukarı doğru projeksiyonlar alır (Şekil 11.5). Muhtemelen, bu yol katekolaminler içeren mavi nokta hücrelerinden ve beyin sapının komşu çekirdeklerinden gelmektedir (Bölüm A.4 ve Şekil A.7).
Yukarıdan aşağıya düzenleyici sistemlerle bağlantılar
beynin daha kaudal kısımlarının aktivitesini düzenlediği ana yol , archistriatum'dan gelir (Zeier ve Karten, 1971; bir inceleme için bkz. Pearson, 1972). Bu yapıda bulunan bazı çekirdeklerin hücrelerinden oldukça büyük bir lif demeti gelir (Şekil 11.6). Bu demete oksipital-mezensefalik yol denir. Hem doğrudan hem de çapraz bileşenleri içerir ve aksonları talamusta, beyin sapının birçok çekirdeğinde ve üst omurilikte sonlanır. Bu sayede, omuriliğin nöronları üzerindeki sinapslar tarafından hareketin doğrudan düzenlenmesi ve bir dizi ara yapı aracılığıyla hareketlerin dolaylı olarak düzenlenmesi gerçekleştirilir. Oksipital-mezensefalik yol görsel çatıdaki bir nöron tabakası ile temas halindedir ve buradan bir efferent-
/

Resim: S.6. Archistriatum'un motor aktiviteyi etkileyen sinyallerin iletilebildiği SMVG ile bağlantıları. Diyagram, güvercin archistriatum'un Zeier ve Karten'e göre alt bölümlere ayrıldığı beş ana çekirdekten dördünü göstermektedir (Zeier ve Karten, 1971). Üç çekirdek, uzun bir inen yola yol açar - oksipito-orta beyin yolu (4). Bu çekirdeklerden biri olan ara çekirdeğin (6) dorsal kısmı , PMIG ile karşılıklı olarak ilişkilidir. Yolun birçok sonlandırma noktasından sadece dördü gösterilmektedir : 1) Şekilde gösterilmeyen retiküler oluşum ile bağlantıların yanı sıra tekto-spinal yol yoluyla motor aktiviteyi etkileyebilen optik çatının derin katmanları ; 2) beyincik (1) ve omurilik üzerine çıkıntı yapan mavi bir nokta (3) g ; 3) muhtemelen retikülo-spinal yolların başladığı pons ve medulla oblongata dahil olmak üzere arka beynin retiküler çekirdekleri (2) ; 4) omuriliğin üst, servikal kısmı . Archistriatum'un alçalan çıkıntıları düz ve çapraz olabilir. 5 - arka kemer; 7 - ara arşistriatum ; 8 - ön arkistriatum.
Bu yapının ny lifleri. Görsel çatı ana görsel alan olmasına rağmen, omurilik dahil görsel işlevi olmayan birçok yapıya yansıtılır . Archistriatum ayrıca orta ve medulla oblongata'nın retiküler oluşumuna aksonlar gönderir, bu da omuriliğe çıkıntı yapar. Oksipital-mezensefalik yolun bazı aksonları mavi noktada son bulur . Bu çekirdeğin nöronları aksonları sadece ön beyine (Bölüm A.4) değil aynı zamanda omuriliğe ve serebelluma da gönderir (Dube ve Parent, 1981; Shiosaka ve diğerleri, 1981). Beyincik, her zamanki gibi, duruş ve hareketin kontrolünde yer alır.
Orta hiperstriatumun dorsal kısmı (Şekil 11.6^ 6) tavuklarda (Bradley ve diğerleri, 1985) ve muhtemelen beç tavuğunda (Bonke ve diğerleri, 1979) PMH ile ilişkilidir. Tavuklarda bağlantılar karşılıklıdır. Zeier ve Karten (Zeier ve Karten, 1971), güvercinde, arkistriatumun bu kısmındaki hafif bir cerrahi yaralanmanın bile oksipitomesensefalik sistemde önemli dejenerasyona yol açtığını bulmuşlardır. Ritchie ve Cohen ( 1977), periektostriat kuşaklardan dorsal arkistriatuma doğru bir çıkıntı buldular (Şekil 11.1b), bu bölgenin görsel bilgi aldığını gösteriyor. Bu gözlemler , arşistriatumun bu bölümünün, görsel uyaranların etkisiyle koşullanan davranışın düzenlenmesinde rol oynayabileceğini göstermektedir. Archistriatum ile görsel çatı arasındaki bağlantıların varlığı da bu sonuçla tutarlıdır. Birçok omurgalıda , görsel çatının, tüm vücut, kafa veya gözlerin hareketlerini gerektirip gerektirmediğine bakılmaksızın, görsel izlemeye dahil olduğu gösterilmiştir (Ingle ve Sprague, 1975; Wurtz ve diğerleri,
.
Anahtarlamalı bir yol aracılığıyla SMVG, ara arşistriatuma da bağlanabilir. PMWG'ye etiketli lösinin eklenmesiyle, beynin yanal bölgesinde etiketli terminallerin alanı tespit edildi (Şekil 11.8). Tavuklarda bu bölümün diğer bağlantıları özel olarak incelenmemiştir. Bununla birlikte, beynin yanal bölgesi, arkistriatumun dorsal yoluna yakındır ve aksonlarını oraya gönderebilir. Bu yol , oksipital-mezensefalik yolun ana kaynağı olan orta arkistriatuma gider .
PMVG, motor aktiviteyi yalnızca arşistriatumun orta kısmıyla olan bağlantılar yoluyla etkilemez. Yükselen paleostriatum (paleostriatum boostatum) ile bağlantıları vardır (Şekil 11.8). Tavukta bu bağlantılar karşılıklıdır. Dorsal arkistriatum durumunda olduğu gibi, karşılaştırmalı anatomi verileri bir varsayımsal işleve işaret etse de, yükseltilmiş paleostriatumun işlevlerine ilişkin deneysel veriler azdır . Embriyonik orijin ve histokimyasal özelliklerdeki benzerlik, yükseltilmiş paleostriatumun memeli bazal ganglionlarının bir kısmına benzer olduğunu düşündürür (Reiner ve diğerleri, 1984 ve s. 325). Bazal gangliyonların işlevine ilişkin ilk belirtiler klinikte elde edildi. Bu yapıların hasar görmesi çok çeşitli hareket bozukluklarına yol açar; ana işlevlerinin yavaş istemli hareketlerin düzenlenmesi olduğu varsayılmaktadır (DeLong, 1974; Kornhuber, 1974). Güvercinlerde, ana bileşeni yükseltilmiş paleostriatum olan paleostriatal kompleksin kimyasal hasarına, hasar yönünde hızlı dönüş, istemsiz hareketler ve postür bozuklukları eşlik eder (Rieke, 1980). Güvercinlerdeki paleostriatal kompleks, öğrenmeyle de ilgili olabilir (Mitchell ve Hali, 1984) ve varsayılan X' bölgesiyle olası özdeşliği daha önce ele alındı (Bölüm 7.3).
Saldırganlık-savunma davranışında ve duyguların ifadesinde yer alan sistemlerle bağlantılar
oksipital-mezensefalik yolun bir parçası olarak gitmez . Güvercinlerde archistriatum'un medial veya posterior çekirdeklerinde hasar görülmedi.
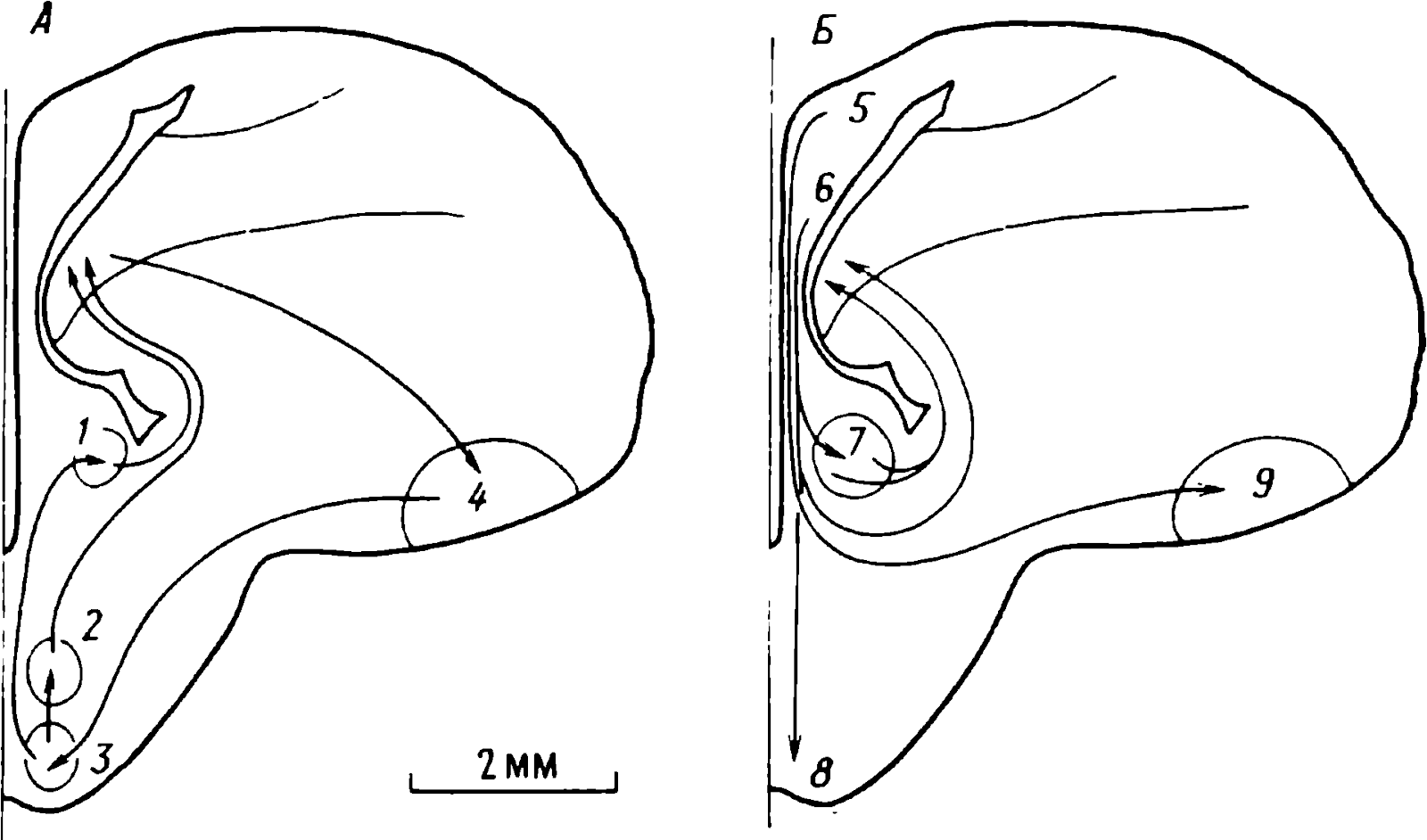
Resim: 11.7. PMVC'nin hipokampal ve hipotalamik bağlantıları. A. Birbirine bağlı yapılar, çeşitli iç organ-endokrin işlevleri düzenleyen bir daire oluşturabilir. B. Hipokampus (6) , lateral (aynı zamanda medial) septal çekirdeklere, hipotalamusa (8) ve PMCG'ye olan çıkıntıları aracılığıyla bu daireye sinyaller enjekte eder . Beynin sadece sol tarafı gösterilmektedir. 1 - lateral septal çekirdek; 2 - talamusun dorsomedial çekirdeği ; 3 - hipotalamusun medial bölgesi; 4 - Archistriatum'un arka bölgesi ; 5 - parahipokampal bölge; 7 - medial ve lateral septal çekirdekler; 9 - Archistriatum'un posteromedial kısmı.
bu yolun liflerinin dejenerasyonuna yol açar. Bu çekirdeklerin projeksiyonları hipotalamusa gider (Zeier ve Karten, 1971). Güvercinlerde, medial hipotalamustan gelen yollar lateral septal çekirdeğe ve talamusun dorso-medial çekirdeğine götürür (Berk, Butler,
. Tavuklarda bu çekirdekler PMVG'ye yansıtılır (Şekil 11.7, L). Buna karşılık, SMVG arşivin arkasına yollar gönderir ve böylece dairesel bağlantıyı kapatır. Hipokampus (Şekil 11.7, B) , septal çekirdekler (Krayniak ve Siegel, 1978a) ve hipotalamus ile dolaylı olarak ve ayrıca SMVG'ye doğrudan bağlantılar yoluyla (Bradley ve diğerleri, 1985) dolaylı olarak bağlanır. Parahipokampal bölge, posterior medial arkistriatuma akson demetleri gönderir ( Krayniak ve Siegel, 1978a).
Beynin PMVG ile ilişkili bazı bölümleri, iç organ ve endokrin işlevlerin düzenlenmesinde ve duygusal ifadenin düzenlenmesinde rol oynar. Diğer pek çok omurgalıda olduğu gibi kuşlarda da hipotalamus, cinsel davranışı ve çevreye verilen diğer endokrin bağımlı tepkileri etkileyen çeşitli düzenleyici işlevlerde yer alır (Hutchison, 1978). Septal bölge ve hipokampus , plazma kortikosteroid seviyeleri üzerinde inhibe edici bir etkiye sahiptir ve bu nedenle strese karşı endokrin tepkilerin modülasyonunda yer alabilir (Bonline ve Bayie, 1973, 1975). Bu sistemin parçaları şek. 11.7, saldırı ve kaçma tepkilerinin organizasyonuna da dahil olabilir (Vowles ve Beazley, 1974). Archistriatum'un medial çekirdeğinin elektriksel olarak uyarılması, tavuklarda bir saldırı tepkisine neden olur (Phillips ve Youngren , 1971). Tersine, medial arkistriatumdaki hasar yaban ördeklerinde kaçınma davranışını azaltır (Phillips, 1964). Phillips ve Youngren (Phillips ve Youngren, 1971), korkuya benzer duygusal tepkilerin tezahürünün , arkistriatumun veya onun efferent liflerinin uyarılmasından kaynaklanabileceğini bulmuşlardır. Anatomik olarak, uyarılan korku ve saldırganlık tepkilerine neden olan alanlar örtüşür, ancak tamamen örtüşmez. Archistriatum bölümlerinin uyarılması veya yok edilmesi , amigdala üzerinde benzer etkilerle memelilerde gözlemlenenlere benzer davranışsal etkilerle birlikte olmuştur (Kaada., 1972). Bu nedenle posterior ve medial çekirdekler, hipotalamusla olan bağlantılarına göre , archistriatum'un “amigdala” kısmı olarak kabul edilirler (Zeier ve Karten, 1971).
memeli amigdalasında olduğu gibi, duygusal davranışın düzenlenmesinde yer alan tek bölge değildir . Çeşitli duygusal davranış türleri , hipotalamus, septal çekirdekler ve hipokampusun uyarılmasıyla değiştirilebilir ( Macphail , 1967; Cannon ve Salzen, 1971; Phillips ve Youngren, 1971; Vowels ve Beazley, 1974). Kuşlarda ve memelilerde septal çekirdekler ile hipokampus arasında karşılıklı bağlantılar vardır. Memelilerde beynin bu bölgelerinin hasar görmesi keşif davranışını, dikkat süresini ve öğrenmeyi bozar ( Gray ve McNaughton tarafından yapılan incelemeye bakın, 1983). Hipokampus ve ilgili yapıların hasar görmesi, hafıza süreçlerini bozar; bu rahatsızlıklar defalarca tarif edilmiştir, ancak mekanizmaları tam olarak anlaşılmamıştır (Bölüm 6.3).
Tavuklarda PMVP ilişkileri, Şekiller 1 ve 2'de özetlenmiştir. 11.8. Aşağıda tartışılacağı gibi talamustan gelen yollar bu alana yaklaşır (Bölüm 11.3) . LMWH'den beynin yanal bölgesine olan projeksiyonlar ve bu alana verilen hasarın tavuklarda damgalama ve pasif kaçınma öğrenme üzerindeki etkilerinden daha önce bahsedilmişti (Bölüm 5.2 ve s. 145). Son zamanlarda, güvercinlerde ilişkisel öğrenme sırasında beynin yanal bölgesine verilen hasarın bazı sonuçları açıklanmıştır. Bir çalışmada, araştırmacılar güvercinleri kırmızı ve yeşil ışığı ayırt etmeleri için eğittiler. Bu alana verilen hasar, bu yeteneği bozmadı (Mogensen, Divae,
. Başka bir çalışmada, kuşlar uyaranları renk tonu, yoğunluk ve desene göre ayırt edecek şekilde eğitildi. Bu durumda hasar öğrenmeyi etkilemiştir (Delius ve diğerleri, 1984). Bu iki çalışmanın sonuçları arasındaki tutarsızlık, kuşların önüne konan görevlerin farklı karmaşıklığı ile açıklanabilir.

Resim: 11.8. SMVG bağlantılarının özet şeması. Afferent yollar düz çizgilerle, götürücü yollar noktalı çizgilerle işaretlenmiştir. Şekilde gösterilen yapıların birçoğu aynı düzlemde yer almamaktadır ve gösterilen yollar gerçek yönlerine tam olarak karşılık gelmemektedir. 1 - Archistriatum'un arka bölgesi; 2 - ara arşistriatumun sırt bölgesi; 3 - beynin yanal bölgesi; 4 — ara hiperstriatum/dorsal hiperstriatum; 5 - ek arşiv; 6 - hipokamp; 7 - neostriatum; 8 - yükselen paleostriatum; 9 - septal çekirdekler; 10 - talamusun dorso-medial bölgesi; Ben - talamusun ventro medial bölgesi .
görevler, ameliyat ve eğitim arasındaki farklı süreler veya diğer bilinmeyen faktörler.
SMVG'nin çeşitli duyusal girdileri vardır, ancak çoğundaki sinyaller bu yapıya girmeden önce kesinlikle önemli bir işlemden geçer. SMVG doğrudan archistriatum'un orta kısmı ile bağlantılıdır. PMSH'nin bu projeksiyon alanı, serebral hemisferlerin inen yollarının önemli bir kaynağıdır (Şekil 11.6). SMVG'den aksesuar hiperstriatuma giden yolların en azından bir kısmının bu yapının görsel olmayan kısmına gitmesi de mümkündür ve bu kısım serebral hemisferlerden gelen yolların ana kaynağıdır. Bu inen yollar aracılığıyla motor aktiviteyi düzenlerler (Karten ve diğerleri, 1973). Benzer bir işlev, PMWG'yi yükseltilmiş paleostriatum ile birleştiren yollar tarafından gerçekleştirilebilir . Tüm bu bağlantılar, postür ve hareketin düzenlenmesinde PMVP'nin yer aldığına işaret etmektedir. Bu alan, arka arkistriatum ve hipotalamus ile olan bağlantıları sayesinde
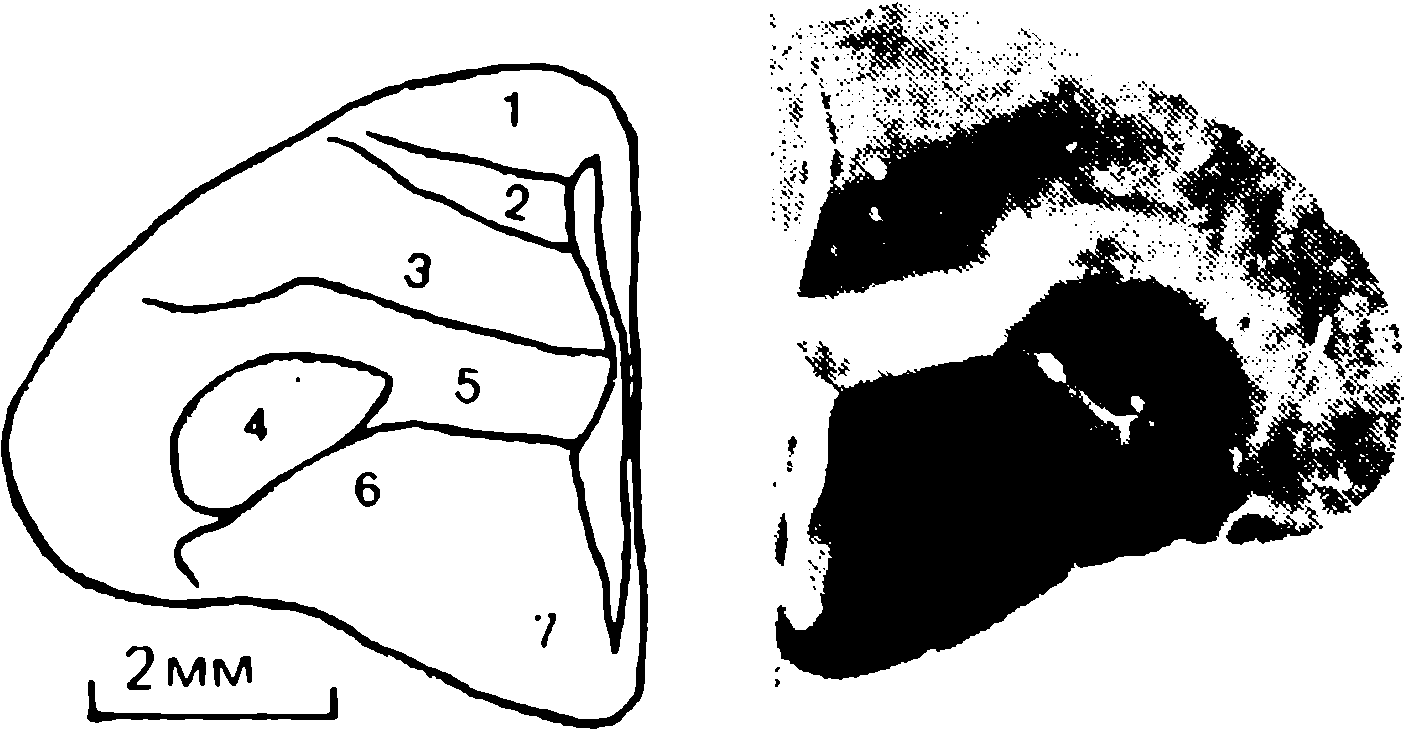
Resim: 11.9. Civciv ön beynindeki asetilkolinesterazın bölgesel dağılımı (2 günlük civcivin beyninden alınan kesit). Asetilkolinesteraz için reaksiyon ürünü, mikrografta koyu görünen hücre gövdelerini ve bunların işlemlerini boyar. Tanımlamalar: 1 — ek hiperstriatum; 2 - dorsal hiperstriatum; 3 - ventral hiper striatum; 4 - ektotriyum; 5 - neostriatum; 6 - yükseltilmiş paleostriatum; 7 - koku alma payı. (McCabe ve diğerleri, 1982.)
(Şekil 11.7) duygusal davranışa da katılabilir. PMVP'nin diğer yapılarla birçok bağlantısı karşılıklıdır ve böyle bir organizasyon , karşılıklı düzenleme yoluyla gerçekleştirilen işlevlerin doğruluğunu artırabilir .
PMVG'nin biyokimyası ve hücresel öğeleri
L Nöroaktif maddeler
PMCG'ye yol açan çeşitli sinir liflerinin çok sayıda aktif madde veya nörotransmitter ve sinir iletimini düzenleyen ve nöromodülatör olarak adlandırılan maddeler salgıladığı varsayılabilir. PMWG'nin içsel nöronları tarafından salgılanan maddeler nedeniyle bu çeşitlilik daha da fazla olabilir.
özelliklere sahip dört biyojenik amin : norepinefrin, dopamin, serotonin ve histamin, ilk üçü PMWH'de bulunur (Davies ve diğerleri, 1983; McCabe ve Hunt, 1983). Bu amino içeren liflerin olası kaynakları, orta beyin ve medulla oblongata'da bulunan hücre gruplarıdır (bkz. Bölüm A.4 ve Şekil A.7).
bölgesel dağılımı bilinmemekle birlikte, güvercin ön beyninde bulunur . Asetilkolini hidrolize eden bir enzim olan asetilkolinesterazın 1-2 günlük civcivlerin beyinlerindeki dağılımı tanımlanmıştır (Şekil 11.9). 6. haftada, yaklaşık olarak aynı kalır. Asetilkolinesteraz, yalnızca Şekil 1'de gösterilen bölgelerde bulunmaz. 11.9, ancak septal çekirdekler dahil diğer bölümlerde de mevcuttur . Memelilerde septal çekirdeklerin asetilkolinesterazın tespit edildiği hücreleri de içerdiğini not etmek ilginçtir (Lewis ve Shute, 1967). Tavuklarda, PMWG dahil olmak üzere ventral hiperstriatumda bulunan enzim. Lekeli hücre gövdeleri bu alanda bulunabilir. Ek olarak, reaksiyon ürünü, sinir liflerinde ve terminallerinde enzimin içeriğini gösteren yaygın olarak dağıtılır . Asetilkolinin PMCH'deki aracılardan biri olduğu varsayımı, esas olarak PMCH içeren sinir dokusunda asetilkolin bağlayan muskarinik reseptörlerin saptanması gerçeğiyle doğrulanır (Bradley ve Noth, 1981). Asetilkolin gerçekten SMVG'de bir nörotransmitter ise, o zaman ya afferent liflerin terminallerinde ya da SMVG'nin dahili nöronal sisteminde üretilir.
Davies ve Horne (Davies ve Horne, 1983), retrograd flüoresan işaretlemeyi ve asetilkolinesterazın histokimyasal tespitini birleştirerek PMCH'ye varsayılan kolinerjik aferent girdinin kaynağını bulmaya çalıştı . Bir günlük tavukların PMWG'sine bir flüoresan işaretleyici eklendi. 20 saat sonra (işaretleyicinin hücre gövdelerine taşınması için gereken süre), beyin izole edildi ve kesitler asetilkolinesteraz için boyandı. Kesitlere parlak ışık altında bakıldığında, asetilkolin esterazın reaksiyon ürününü içeren hücre gövdeleri görülebiliyordu. Bir ultraviyole ışık kaynağına geçerek, bu hücre gövdelerinin floresan olup olmadığını belirlemek mümkün oldu. Bazı telensefalik alanlarda, aynı perikaryada floresan bir etiket ve asetilkolinesteraz için kahverengi bir reaksiyon ürünü tespit edildi. Bu nedenle, bu nöronların SMVG'ye yansıtılması mümkündür; dağılımları Şek. 11.10. Bu tür liflerin ana kaynakları septal çekirdekler ve hipokampus olup, bağlantılar iki taraflıdır. İpsilateral neostriatum ve ventral hiperstriatumun kaudal kısmı da PMWG'ye asetilkolinesteraz içeren lifler gönderir. Damgalama sırasında asetilkolinesteraz aktivitesinde geçici bir artış olduğu bulunduğundan (Haywood ve diğerleri, 1975), bu afferent yolakların damgalama sürecine dahil olması muhtemeldir.
PMVG'de başka nöroaktif maddeler de tanımlanmıştır. Böylece hücre gövdeleri metiyonin-enkefalin içerir ve sinir lifleri kolesistokinin içerir (McCabe, Hunt, 1983). Bu peptitler oldukça aktiftir ve doğrudan nöronlara uygulandığında onları uyarabilir, inhibe edebilir veya birleşik bir etki sergileyebilir. PMGB'de inhibe edici nöronlarda bulunan bir aracı olan γ-aminobütirik asidin varlığına dair veri yoktur .
Yukarıdaki açıklamadan, PMWG'de bulunan nöroaktif maddeler hakkındaki bilginin parçalı kaldığı açıktır.
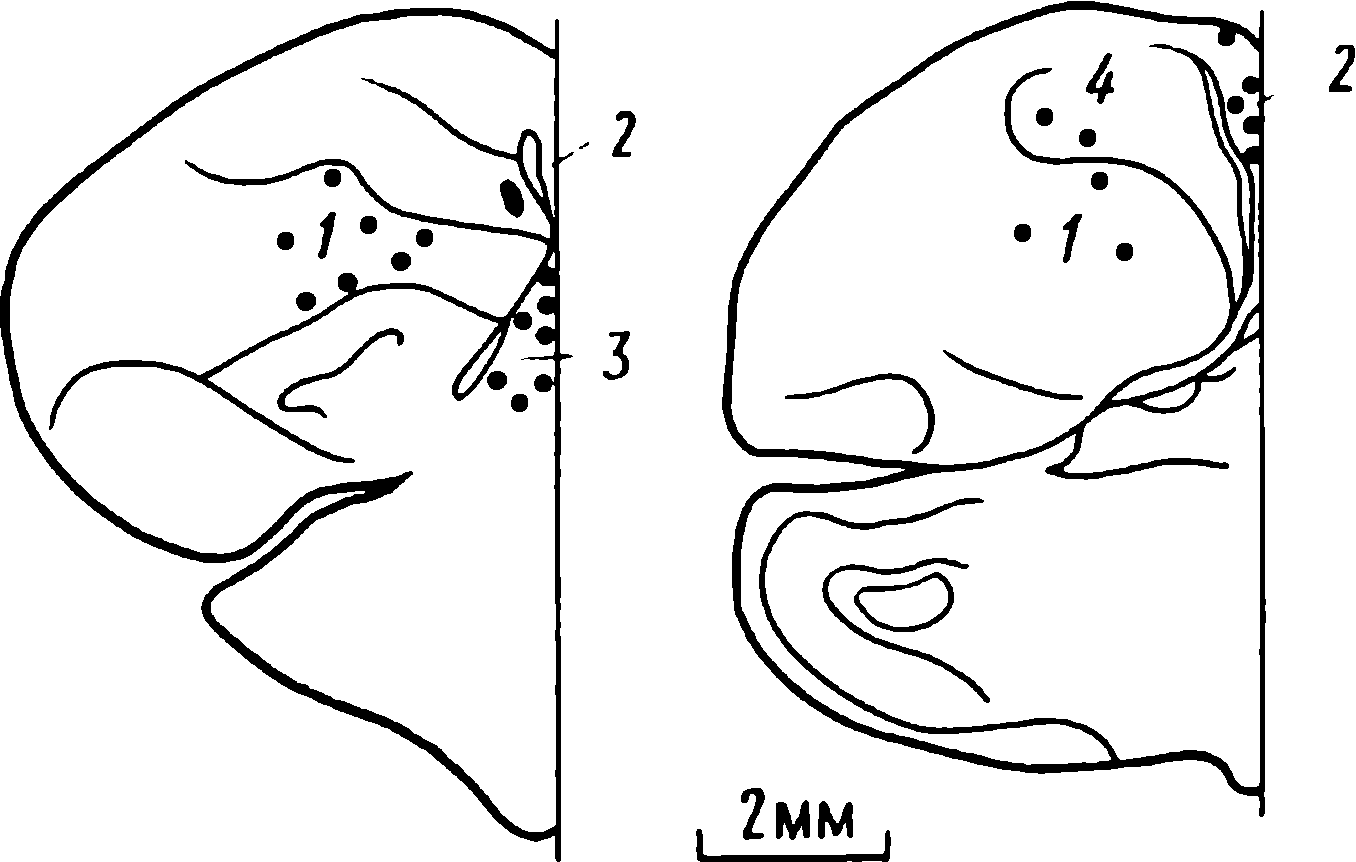
Resim: 11.10. Aksonları PMCH'ye gönderen asetilkolinesteraz içeren perikaryaların lokalizasyonu. Floresan işaretleyicinin PMIG'e giriş yeri siyah bir oval ile işaretlenmiştir. Çift etiketli perikaryaların lokalizasyonu siyah noktalarla belirtilmiştir. Soldaki diyagram, sağdakinden daha rostral bir bölgeyi temsil ediyor. 1 - neostriatum; 2 - hipokamp; 3 - septal çekirdekler; 4 - ventral hiperstriatum. (Davies, Noth, 1983.)
bu nöroaktif maddeler hakkında çok daha fazlasının bilinmesi gerekiyor . Özellikle bu maddeleri içeren sinir liflerinin kaynaklarını, bu liflerin hedeflerini ve SMVG içindeki sinaptik organizasyonlarını bilmek önemlidir.
Hücresel organizasyon
PMVG'nin hücre mimarisi hakkında çok az şey bilinmektedir. Dokunun gümüş nitrat ile emprenye edildiği Golgi yöntemi kullanılarak , nöronlar ve bazı süreçleri açıkça görülebilir. Bradley ve Horne (Bradley ve Noth, 1982), SMVG'deki sinir hücrelerinin, memelilerin optik çatısında veya neokorteksinde nasıl konumlandıklarına benzer şekilde katmanlar halinde düzenlendiğini buldular. PMVG'deki sinir hücreleri , perikaryonun boyutuna ve şekline göre sınıflandırılabilir. Bu kriterlere göre 4 tip hücre tanımlanmıştır. Bunlardan üçü Şekil l'de gösterilmiştir. 11.11. Piramidal gövdeli hücrelerin aksonları (Şekil 11.11, C ) , PMSH'den ayrılmadan önce genellikle iki dala ayrılır. Bu hücreler neredeyse kesinlikle projeksiyon nöronlarıdır. Aksonları genellikle PMIG dışında izlenemediğinden, diğer hücrelerin de projeksiyon hücreleri olup olmadığı net değildir. Hücrelerin yaklaşık dörtte biri küçük piramidal veya yıldız şeklinde perikaryaya sahipti; incirde. 11.11 gösterilmezler. Bu hücrelerin işlemlerinin tamamen PMVG içinde olması muhtemeldir. Bu nöronları benzer şekilli memeli omurilik nöronlarıyla karşılaştırmak
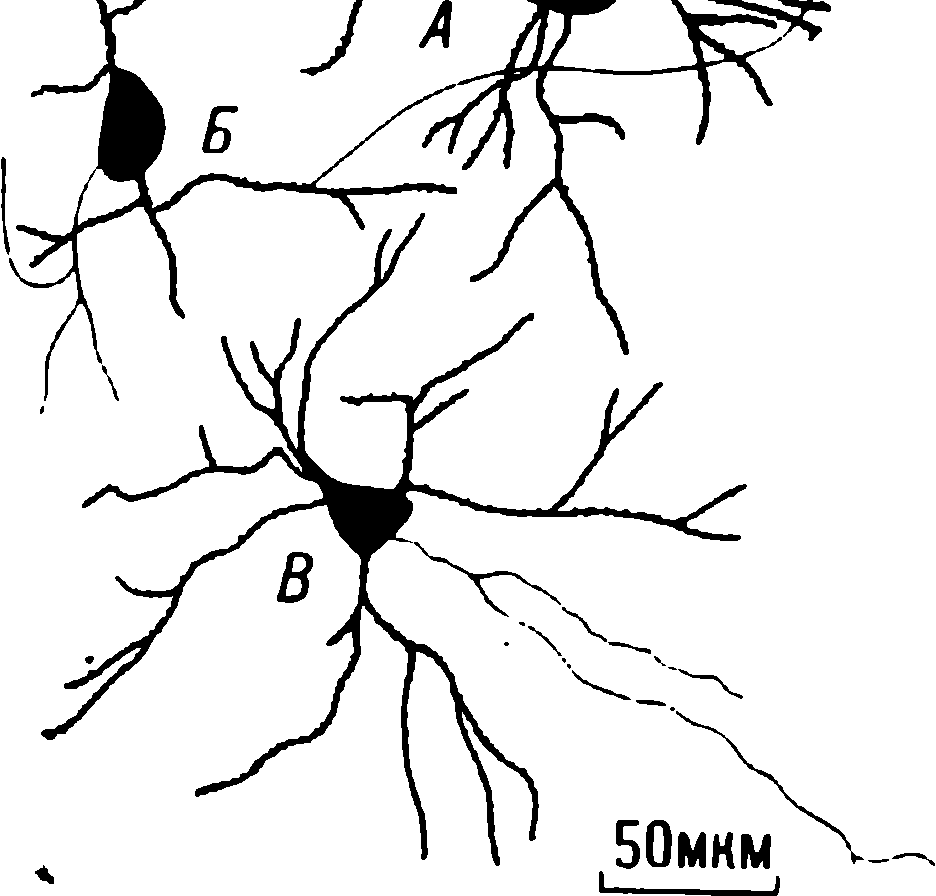
Resim: 11.11. Bir çizim aparatı kullanılarak SMVG'nin nöronlarının çizilmesi. Toplamda 140 Golgi emdirilmiş hücre analiz edildi. Işıkta bekletilen bir günlük civcivlerin beyinlerinden kesitler alındı. Dendritler, dikenlerin varlığı ile tanımlanabilir, aksonlar düzgün konturlara sahiptir. A. 56 nöron (%40) büyüktü ve çok sayıda birincil dendritleri vardı. Aksonların seyri oldukça değişkendi. B. 21 nöron (%15) büyük oval gövdelere sahipti. Bu hücrelerin aksonları, hücre gövdesinin yakınında birkaç dala bölünmüştür. B. 27 nöron (%20) büyük piramidal gövdelere sahipti. Bu hücrelerin aksonları genellikle iki dala bölünerek PMCH'nin ötesine geçer. (Bradley, Noth, 1982.)
ve boyutlarına bakıldığında, en azından bir kısmının inhibitör işlevi olan hücreler arasında yer aldığı söylenebilir.
Şu anda, ışık mikroskobu ile tespit edilen PMIG'nin sito-mimari yapısını bu alandaki aracıların ince yapısı ve kimyası ile ilişkilendirmek gereklidir. Bu tür bilgilerle, sinapsların organizasyonu ve yerleşimi hakkında nicel bilgilerin yanı sıra PMCG nöronlarının elektrofizyolojisine ilişkin verilerle, beynin bu bölgesinin işleyiş ilkelerini daha iyi anlamak mümkün olacaktır.
Yorumlar ve akıl yürütme: kuşlarda ve memelilerde sinir yolları
PMVC'ye giden görsel yollar göz önüne alındığında, maymun korteksinin inferotemporal bölgesinden bahsedeceğiz, çünkü bu bölgelerin her ikisi de multisinaptik ve oligosinaptik yollardan görsel sinyaller alıyor. Bu benzetme yararlı olabilir , ancak neden abartılmaması gerektiğini belirttik. Bununla birlikte, artık PMCH'nin bağlantıları hakkında daha fazla şey bildiğimize göre, primat beyninin herhangi bir bölümünün işlevsel veya anatomik olarak PMCH'ye karşılık gelip gelmediği sorusuna geri dönebiliriz . Bu tür karşılaştırmaların zorluğu bizim tarafımızdan Sec. S.2. Bununla birlikte, tavuklarda PMVP'ye verilen hasarın ve insanlarda ve diğer primatlarda belirli beyin yapılarına verilen hasarın benzer şekilde hafıza süreçlerini etkilediğini daha önce belirtmiştik (Bölüm 6.3 ve 6.4). PMCG'nin bu primat beyin yapılarına benzer olduğuna inanmak için herhangi bir neden var mı? Bu soruyu cevaplamaya çalışırken, bu tür karşılaştırmaların dayandığı kırılgan temeli akılda tutmak gerekir . Memeli merkezi sinir sistemi , diğer herhangi bir omurgalı sınıfından çok daha yoğun bir şekilde incelenmiştir. Memeli beyni hakkındaki bilgimizle karşılaştırıldığında , kuş beyni hakkındaki anlayışımız neredeyse ilkel. Bu tutarsızlık tek başına herhangi bir karşılaştırmayı zorlaştırır ve bu zorluk, her sınıf içindeki büyük tür farklılıklarının varlığıyla daha da artar. Sonuç olarak , yalnızca oldukça genel karşılaştırmalar geçerli kabul edilebilir. Yapının embriyonik kökeninin benzerliği, diğer bölümlerle bağlantıları ve fizyolojisi esas alınmalıdır .
PMVG'nin beynin hangi bölümlerine benzemediğini söylemek güvenlidir . Memelilerin birincil duyusal kortikal bölgelerine benzemez. Nadir istisnalar dışında bu alanlar, PMCH'nin özelliği olmayan talamusun projeksiyon çekirdeklerinden spesifik duyusal yollar alır. PMVG'ye ve hipokampus ve dentat girustan oluşan eski memeli korteksine benzemez: embriyogenezdeki hipokampus nöral tüpün medial duvarından oluşurken ventral hiperstriatum lateral duvardan kaynaklanır (Şekil A. 4, B). Bununla birlikte, hipokampus PMCH'ye yansıtılır ve bu ilişki çok şey anlatır. Arama alanı, PMCG'nin talamusun medial çekirdeklerinden doğrudan girdi alması gerçeğiyle daha da daraltılır (Şekil 11.8). Primat serebral korteksinin iki bölgesi , prefrontal ve cingular, talamusun medial çekirdeklerinden afferent yollar alır ve hipokampusla güçlü bağlantıları vardır (bkz . Brodal tarafından yapılan inceleme, 1981).
Prefrontal korteks, frontal lobun ön kısmının yüzeyini kaplar. Bu alanın bağlantıları, PMWG bağlantılarının analiz edildiği sırayla ele alınacaktır. Prefrontal korteksin bazı bağlantıları Şekil 1'de gösterilmektedir. 11.12. 1) Prefrontal korteks, talamusun duyusal röle çekirdeklerinden gelen yolları almaz . Bununla birlikte, duyusal bilgi ona birincil duyusal alanlara bitişik kortikal alanlardan ulaşır, yani bilgi son kısımdan prefrontal kortekse girer.
140
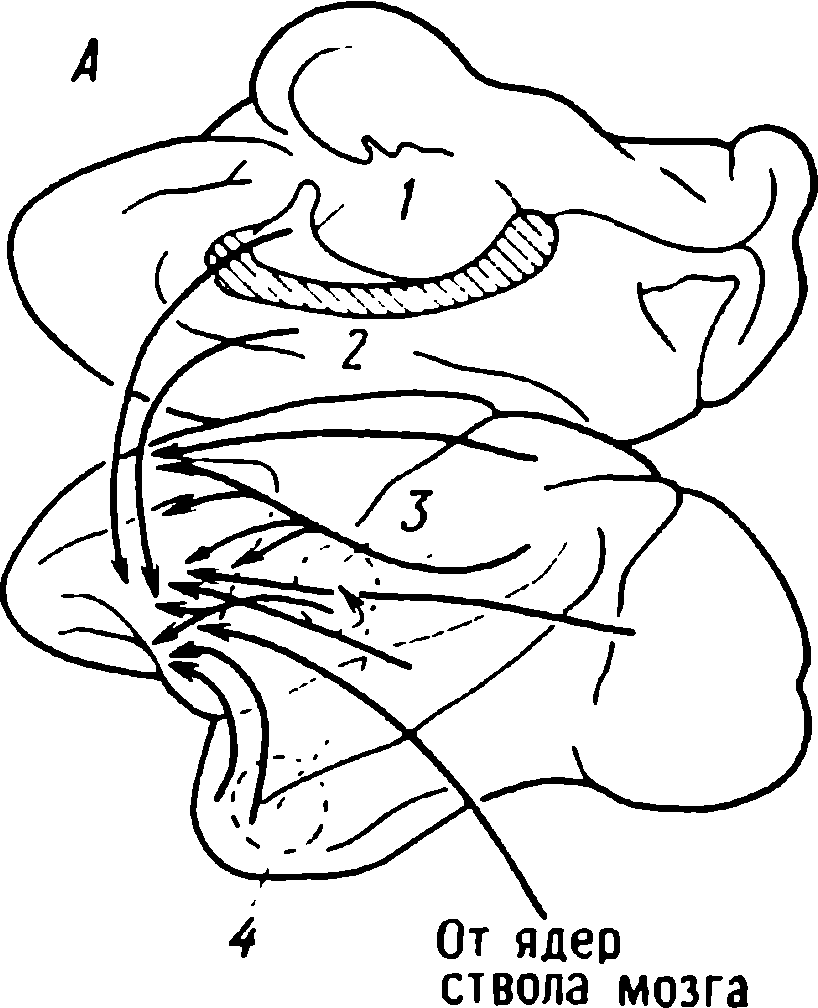
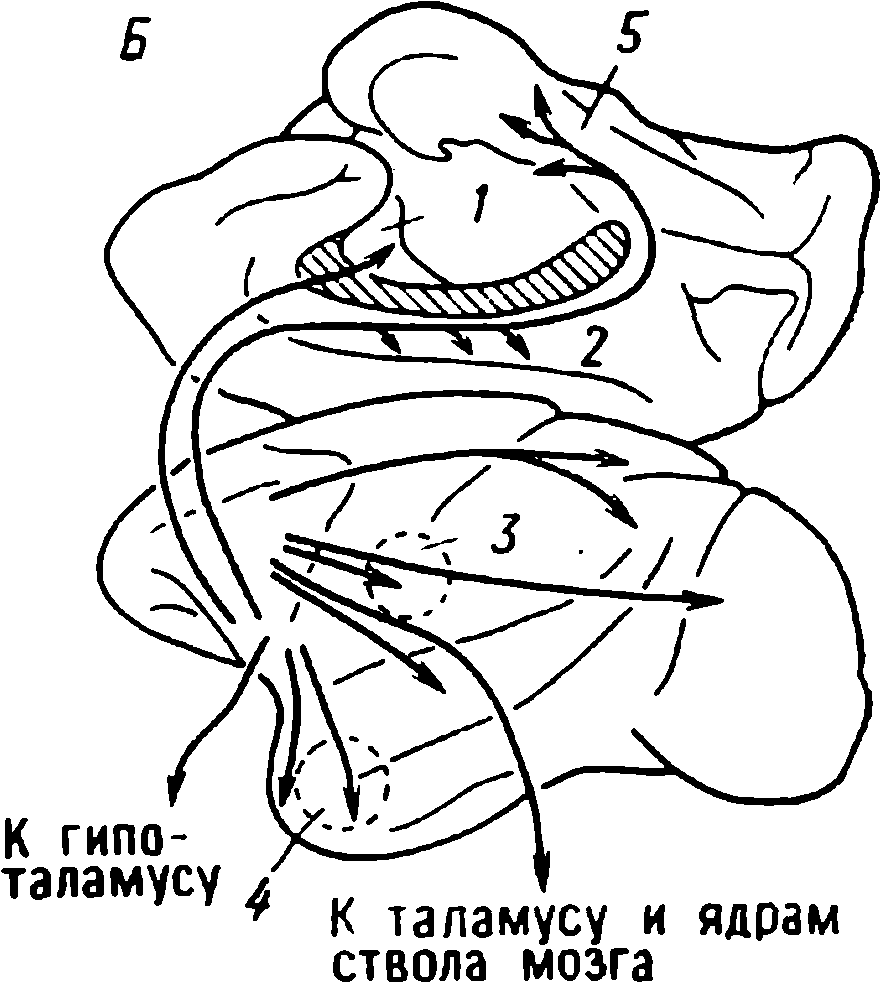
beyin. SMVG, benzer bir duyusal girdi organizasyonuna sahiptir: birkaç görsel yoldan biri dışında, SMVG'ye giden tüm duyusal yollar, ona telensefalonun diğer bölümlerinden gider. 2) Prefrontal korteks, sinir liflerini doğrudan serebral korteksin ana motor yoluna - piramidal yola göndermez. Ancak oluşumunda yer alan parietal loba yansıtılır (Russell ve DeMeyer , 1961). Prefrontal korteks ayrıca talamusa, retiküler formasyona ve locus coeruleus'a da çıkıntı yapar. SMVG, ön beyin dışında uzanan yapılarla doğrudan bağlantılı değildir , ancak dorsal ve muhtemelen orta arkistriatum yoluyla dolaylı olarak onlara bağlanır. Prefrontal korteks, bazal ganglionlara bağlantılar gönderir; PMVG, genellikle prefrontal korteksten bağlantıları alan bazal ganglionların o kısmıyla karşılaştırılan yükselmiş paleostriatum ile ilişkilidir . 3) Amigdala, hipotalamus ve talamusun dorso-medial çekirdeğinin prefrontal kortekse çıkıntıları vardır. Posterior ve medial arkistiyal atum amigdalaya karşılık geliyorsa, bu prefrontal korteks bağlantıları, Şekil 1'de gösterilen PMCG bağlantılarına çok benzer.
C. 4) Prefrontal korteks , ek anahtarlar aracılığıyla hipokampa bağlanır. Bu yapının aksonları, sırayla talamusa uzanan septal çekirdeklere gider. Talamusun dorso-medial çekirdeğindeki hücreler aksonları prefrontal kortekse gönderir. Tavuklarda, hipokampus doğrudan ve dorsal septal çekirdekler aracılığıyla SMVG'ye bağlantılar gönderir . Bu projeksiyon kolinerjik olabilir ( Davies & Noth, 1983). Prefrontal korteks, ventral septal bölgedeki yapılardan kolinerjik yolları alır (Domesick, 1976; Meibach ve Siegel, 1977). Bu sistem , şiddetli bir amnezinin olduğu Alzheimer tipi yaşlılık psikozlu hastalarda dejenere olur (Davies ve Maloney, 1976; Whitehouse ve diğerleri, 1982; Coyle ve diğerleri, 1983; Mountjoy ve diğerleri, 1984). . 5) Prefrontal korteks, afferent bağlantıları aldığı bölümlerle karşılıklı olarak bağlantılıdır. Aynı durum PMVG için de geçerlidir.
PMVC ve cingular korteks arasındaki benzerlik daha az belirgindir, ancak yine de açıklamayı hak ediyor. SMVG , talamusun medial yerleşimli dorso-medial ve ventromedial çekirdeklerinden bağlantılar alır . Singular korteks medial yerleşimli antero-medial ve dorsomedial talamik çekirdeklerden bağlantılar alır. Primatlarda, arkikorteksin septal bölgesi ve subiküler bölgesi, cingular korteks ile ilişkilidir . Alt grup, tavukların para hipokampal bölgesine benzer olarak düşünülebilir (Benowitz ve Karten, 1976; Krayniak ve Siegel, 1978a). Bu bölgenin SMCH'ye doğrudan yolları olmamasına rağmen, hipokampus ve septal çekirdekler yoluyla ona bağlanabilir (Krayniak ve Siegel, 1978a; Bradley ve diğerleri, 1985). Primatlarda, cingular korteks, prefrontal korteks de dahil olmak üzere afferent yolları aldığı bir dizi yapıya bağlantılar gönderir .
Tabii ki, tavuklardaki PMCG ile primatların prefrontal ve cingular korteksi arasındaki bağlantılar birbirinden önemli ölçüde farklıdır ve hatta görünüşteki benzerlik yüzeysel olabilir. Örneğin, güvercin ve sıçandaki dorso-medial çekirdek, septal bölgenin ventral kısmından bağlantıları eşit olarak alır (Krayniak ve Siegel, 1978b; Meibach ve Siegel, 1977); Aynı zamanda güvercinde bu çekirdek hipotalamustan yollar alırken (Berk ve Butler, 1981), memelilerde böyle bir bağlantıya dair veri yoktur (Raisman, 1966 ). Bununla birlikte, PMIG ile serebral korteksin açıklanan bölgeleri arasında, hafıza süreçlerindeki olası katılımlarının karşılaştırılmasına izin verecek kadar benzerlik vardır.
İnsanlarda anterior medial talamusun hasar görmesi şiddetli amneziye neden olur (Briegley, 1977). Talamusun bu kısmı, hipokampus ve septal çekirdekler dahil olmak üzere birçok yapıdan bağlantı alır. Talamusun anterior ve medial çekirdekleri, septal bölge ve hipotalamusta yer alan mamiller çekirdekler aracılığıyla, hipokampustan ve ona bitişik subikulumdan sinyaller alır. Bu yapılardan herhangi birinin hasar görmesi amneziye neden olur . Prefrontal ve singular korteks, bu yapılardan doğrudan veya anahtarlanmış yolların varış noktalarıdır. Warrington ve Weiskrantz ( 1982) , hipokampusun , prefrontal bölgeyi içeren frontal kortekste iz oluşumunu kontrol edebileceğini ileri sürmüştür. Griffith (Griffith, 1966) hipokampus ve neokorteks arasındaki etkileşime de işaret etti. Hafıza izlerinin temeli olarak gördüğü sinaptik değişikliklerin neokortekste gerçekleştiğine ve bu sinaptik değişikliklerin hipokampal nöronlardan gelen sinyaller tarafından kontrol edildiğine inanıyor. Squire ve diğerleri ( 1984) ve Milner (Milner , 1987) de hipokampusu içeren medial temporal lob yapılarının neokorteks ile etkileşime girdiğine inanırlar. Bu etkileşimin bir sonucu olarak , neokorteksteki varsayılan nöral değişiklikler, nispeten uzun bir süre boyunca kademeli olarak pekiştirilir . Böyle bir etkileşime bağlı bellek izleri medial temporal lobun hasar görmesi ile oluşamaz. Üstelik hasar anında konsolidasyon sürecinde olan izler parçalanır. Sonuç olarak, yaralanmadan birkaç yıl önce meydana gelen olaylar için bir hafıza kaybı vardır .
Bu model doğru olsun ya da olmasın, insanlarda ve diğer memelilerde birbirine bağlı birkaç beyin bölgesinin hafıza için kritik olan bir sistem oluşturduğuna dair güçlü kanıtlar vardır (bkz. Vinogradova, 1975; Weiskrantz, 1982). Bireysel bileşenlerinin tam işlevi tam olarak açık değildir, ancak bellek için önemleri inkar edilemez. Piliçlerde, sol ve sağ PMVP'nin bellekteki rolleri farklı olmasına rağmen, PMVP de benzer işlevlere sahiptir (Bölüm 7). Krushinskaya'nın çalışmasında (Vinogradov'dan alıntılanmıştır, 1978), kuşlarda hipokampusun da hafızaya katılabileceğinin bir göstergesi vardır. Krushinskaya , kilerde fındık depolayan Nucifraga caryocatases'i inceledi . Hipokampusa verilen hasardan sonra kuşlar, kilerden 3 saatten fazla izole edilirlerse kilerlerin yerini unuturlar.Sahgal ( 1984) , hipokampusa verilen hasarın güvercinlerde tanıma üzerindeki etkisini inceledi . Kuşlara bir uyaran verildi ve birkaç saniye sonra ya aynı ya da farklı bir uyaran gösterildi. Kuşlar, 1. ve 2. uyaranlar aynıysa bir kaldıracı, farklıysa başka bir kaldıracı gagalamak zorunda kaldı. Hipokampusta hasar olan güvercinlerde bu görevin çözümü bozulmuştur.
Kuşların ve memelilerin beyninin organizasyonunda belirli bir benzerlik vardır, ancak aynı zamanda önemli temel farklılıklar da vardır. Bu iki omurgalı türünün beyinleri embriyonik olarak oldukça benzer görünen nöral tüplerden gelişse de , memeli beyninin serebral hemisferlerinin iç yapısı kuşunkinden çok farklıdır. Bu nedenle, kuşların ve memelilerin beyinlerindeki bazı yapıların yukarıda anlatılan benzerliğinin, tesadüfen ortaya çıkmış olması muhtemeldir. Bununla birlikte, kuşları ve memelileri ortak bir atadan ayıran devasa zaman aralığı göz önüne alındığında, bu benzerlik hala şaşırtıcı. Evrim sürecinde, belirli bilgi türlerinin analizi, depolanması ve bellekten alınması için en verimli olan bir sinir organizasyonu seçilebilirdi . Tabii ki, genel temelin varyasyonları olmaması olamaz. Yukarıda açıklanan sistemlerin organizasyon detaylarındaki farklılık şaşırtıcı değildir.
Özet
PMCG'nin nöronları, memeli serebral korteks hücrelerinin aksine, katmanlar halinde organize edilmemiştir. PMWG'de farklı hücre türleri vardır. Bazıları ara nöron gibi görünürken, diğerleri beynin diğer bölgelerine aksonlar gönderir. PMVG, aracı ve modülatör olarak işlev görebilen önemli sayıda nöroaktif maddeyi ortaya çıkardı . PMVG, çoğu duyusal sistemden bağlantı alır ve olası bir istisna dışında, bu alana giren bilgi, önbeynin diğer duyusal alanlarında önceden işlenir. Doğrudan veya anahtarlamalı yollar aracılığıyla SMVG'nin ön beyinden bir veya iki ana çıkışa erişimi vardır. Bu bağlantılar aracılığıyla PMVP, duruş, hareket ve görsel olarak yönlendirilen davranış üzerinde düzenleyici bir etki gösterebilir. PMVP'nin ayrıca saldırı, kaçınma ve iç organ-endokrin işlevleri gibi davranışları düzenlemeye dahil olabilecek sistemlerle bağlantıları vardır. PMVG bağlantıları, primat serebral korteksin prefrontal ve cingular bölgelerine benzer. PMHG ve neokorteksin belirtilen bölümlerini içeren sistemlerin benzerliği, işlevlerinin hafıza ile ilişkili olmasıyla da ifade edilir.
EDEBİYAT
Adamo NJ Tavuk, kuzgun ve Afrika cennet papağanı, J. comp. Neurol., 1967, 131, 337-356.
aenowitz L. L, Karten HJ Güvercin parahipokampal bölgesine traktus infundibuli ve diğer afferentler, Brain Res., 1976, 102, 174-180.
Bentivoglio M., Kuypers HGJ M Catsman-Berrevoets CE, Loewe H., Dann 0. Taşıyıcı olan iki yeni floresan retrograd nöronal izleyici
uzun mesafelerde ted, Neurosci. Lett., 1980, 18, 25-30.
Berk ML, Butler AB Güvercinlerde medial preoptik çekirdeğin ve medial hipotalamusun efferent projeksiyonları, J. comp. Neurol., 1981, 203, 379-399.
Bonke BA, Bonke D., Scheich H. Gine tavuğundaki (Numida meleagris) işitsel ön beyin çekirdeklerinin bağlantısı . Celi Tissue Res., 1979, 200, 101-121.
Bouille C., Bayie J. D. Güvercinlerde bazal ve stres kaynaklı hipotalamik-hipofiz-adrenokortikal aktivite üzerindeki limbik uyarım veya lezyonun etkileri, Neuroendocrinology, 1973, 13, 264-277.
Bouille C., Bayie JD Kuşlarda bazal hipofiz adrenokortikal fonksiyon üzerinde septal çekirdeklerin etkisi, Neuroendocrinology, 1975, 18, 281-289.
Bradley P., Horn G. Damgalama: civciv beyninin bölümlerindeki kolinerjik reseptör bölgeleri üzerine bir çalışma, Exp. Brain Res., 1981, 41, 121-123.
Bradley P., Horn G. Civcivdeki hyperstriatum ventrale'nin Golgy analizi, J. Anat, 1982, 134, 599-600.
Bradley P., Davies D, C., Horn G. Evcil civcivin (Gallus domesticus) hyperstriatum ventrale bağlantıları, J. Anat., 1985, 140, 577-589.
Brazier M. A. B. Anestezinin beyin üzerindeki bazı etkileri, Br. J. Anaesth., 1961, 33, 194-204.
Brierley JB Amnezik durumların Nöropatolojisi. In: Amnesia (editörler CWM Whitty ve OL Zangwill), 1977, s. 199-223. Butterworth, Londra.
Brodal A. Klinik tıpla ilgili olarak nörolojik anatomi, 1981, Oxford University Press, New York.
Brown MW, Horn G, Anestezi uygulanmış ve anestezi uygulanmamış kedilerin hipokampal bölgesindeki nöronların duyu yollarının uyarılmasına tepkisi, Brain Res., 1977, 123, 241-259.
Brown MW, Horn G. Eğitimli maymunların hipokampal bölgesinden kaydedilen bağlama bağlı nöronal tepkiler, J. Physiol., Londra., 1978, 282, 15-16P.
Brown MW, Horn G. Civciv beynindeki nöronal plastisite: görsel deneyimin hiperstriatal nöronlar üzerindeki elektrofizyolojik etkileri, Brain Res., 1979, 162, 142-147.
Cannon RE, Salzen EA Yumurtadan yeni çıkmış civcivlerde beyin uyarımı, Anim, Behav., 1971, 19, 375-385.
Clutton-Brock J. Anestezik ajanların merkezi sinir etkilerinin önemi, Br. J. Anaesth., 1961, 33, 214-218.
Coyle J, T., Price DL, DeLong MR Alzheimer hastalığı: bir kortikal kolinerjik innervasyon bozukluğu, Science, 1983, 219, 1184-1190.
Davies DC, Horn G. Piliç hyperstriatum ventrale'nin varsayılan kolinerjik afferentleri: kombine bir asetilkolinesteraz ve retrograd floresan etiketleme çalışması, Neurosci. Lett., 1983, 38, 103-107.
Davies DC, Horn G., McCabe BJ Evcil civcivlerde telensefalik katekolamin seviyelerindeki değişiklikler. Yaş ve görsel deneyimin etkileri, Devi. Brain Res., 1983, 10, 251-255.
Davies DC, Horn G., McCabe BJ Noradrenalin ve öğrenme: nörotoksin DSP4'ün evcil civciv Behav'da damgalama üzerindeki etkileri. Neurosci., 1985, 99, 652-660.
Davies P., Maloney AJF Alzheimer hastalığında Merkezi kolinerjik nöronların seçici kaybı, Lancet, 2, 1976, 1403.
Delius JD, Bennetto K. Kuş ön beynine deri duyusal projeksiyonlar. Brain Res., 1972, 37, 205-221.
Delius JD, Runge TE, Oeckinghaus H. Güvercin ön telensefalonuna kısa gecikmeli işitsel projeksiyon, Exp. Neurol., 1979, 63, 594-609.
deli /. D., Jâger R., Frisel M. Güvercinlerde lateral telensefalik lezyonlar, Behav'da görsel ayrımcılığı etkiler. Brain Res., 1984, 11, 249-258.
DeLong MR Bazal ganglionların motor fonksiyonları: hareket sırasında tek üniteli aktivite. İçinde: Nörobilimler üçüncü çalışma programı (eds. F. O. Schmitt ve F. G. Worden), 1974, s. 319-326, MIT Press, Cambridge, Mass.
Broca diyagonal bandının çekirdeğinin Domesick V. B. Projectioris'i, Anat. Rec., 1976, 184, 391-392.
Dube L., Ebeveyn A. Kuş beynindeki monoamin içeren nöronlar: 1. Flüoresan ve asetilkolinesteraz histokimyası yoluyla civcivin (Gallus domesticus) beyin sapı üzerine bir çalışma, J. comp. Neurol., 1981, 196, 695-708.
Erulkar SD Güvercin beynindeki dokunsal ve işitsel alanlar, J. komp. Neurol., 1955, 103, 421-457.
F4lix B., Kesar S., Roesch, R. Landes kaz, Exp. Brain Res., 1983, 53, 173-182.
Tavuk ve diğer kuşların nazik MJ Tat verici davranışı. İçinde: Kuşlarda davranışın sinirsel ve endokrin yönleri (eds. P. Wright, PG Caryl ve DM Vowles), 1975, s. 305-318, Elsevier, Amsterdam.
Gottlieb G. Kuşlarda tür tanımlamasının geliştirilmesi. 1971 Üniversite Chicago Press , Şikago.
Gottlieb G,, Klopfer PH Gelişimsel yaşın işitsel ve görsel damgalama ile ilişkisi, J. comp. fizik. Psychol., 1962, 55, 821-826.
Gray JA, McNaughton N. Septal ve hipokampal lezyonların davranışsal etkilerinin karşılaştırılması: bir gözden geçirme. nörobilim Biyodavranış. Rev., 1983, 7, 119-188.
Griffith JS Belleğin doğasına ilişkin bir teori, Nature, Londra., 1966, 211, 1160-1163.
Brüt CG Alttemporal korteksin görsel işlevleri. İçinde: Duyusal fizyolojinin el kitabı (ed. R. Lung), 1973, VII, 3B, s. 229-238, Springer, W. Berlin.
Brüt CG, Bender DB, Rocfia-Miranda CE Inferotemporal korteks: tek birim analizi. İçinde: Nörobilimler üçüncü çalışma programı (eds. F. O. Schmitt ve F. G. Worden), 1974, s. 229-238, MIT Press, Cambridge, Mass.
Guselnikov V.I., Morenkov ED, Do Kong Hun. Güvercin hiperstriatumunun görsel projeksiyon bölgesindeki nöronların alıcı alanlarının reaksiyonları ve özellikleri . Nörofizyoloji, 1976, 8, 230-236.
Haywood J., Hambley J., Rose S. Civciv beyninde asetilkolin metabolizmasında yer alan enzimlerin aktivitesi üzerinde baskılayıcı bir uyarana maruz kalmanın etkileri, Brain Res., 1975, 92, 219-225.
Horn G., Wiesenfeld Z. Kedide dikkatli davranış: elektrofizyolojik çalışmalar, Exp. Brain Res., 1974, 21, 67-82.
Horn G., McCabe BJ, Bateson PPG Damgalamadan sonra civciv beyninin otoradyografik bir çalışması, Brain Res., 1979, 168, 361-373.
Hubel D.H. Exploration of theprimer görsel korteks, 1955-1978, Nature, Londra., 1982, 299, 515-524.
Hutchison JB (ed.) Cinsel davranışın biyolojik belirleyicileri. John Wiley, Chichester, 1978.
Ingle D., Sprague J. (editörler). Orta beyin tektumunun sensorimotor işlevi, Neuro sci. Res. Program Bülteni, 1975, 13, 169-288.
Kaada BR Fonksiyonel temsillere referansla amigdaloid kompleksinin stimülasyonu ve bölgesel ablasyonu. İçinde: Amigdalanın nörobiyolojisi (ed. BE Elftheriou), 1972, s. 205-281, Plenum Press, New York.
Karten H. J „ Hodos W., Nauta WJH, Revzin AM Kuş telensefalonunun “görsel Wulst”unun sinirsel bağlantıları. Güvercin (Columba livia) ve baykuşta (Speotyto cunicularla) deneysel çalışmalar , J. comp. Neurol., 1973, 150, 253-278.
Kornhuber HH Ceçebral korteks, serebellum ve bazal gangliyonlar: motor fonksiyonlarına giriş. İçinde: Nörobilimler üçüncü çalışma programı (eds. F. O. Schmitt ve F. G. Worden), 1974, s. 267-280, MIT Press, Cambridge, Mass.
Krayniak PF, Siegel A. Güvercindeki hipokampus ve bitişik bölgelerin etkili bağlantıları, Brain Behav. Evol., 1978a, 15, 372-388.
Krayniak P, F „ Siegel A. Güvercin septal alanının efferent bağlantıları, Brain Behav. Evol., 1978b, 15, 389-404.
Kristensson K., Olsson Y. Proteinin retrograd aksonal taşınması, Brain Res., 1971, 29, 363-365.
LaVail JH, LaVail MM Merkezi sinir sisteminde retrograd aksonal taşıma, Science, 1972, 176, 1416-1417.
Lewis PR, Shute CCD Kolinerjik limbik sistem: hipokampal formasyona, medial kortekse, yükselen kolinerjik retiküler sistemin çekirdeklerine ve subfornikal organa ve supra-optik tepeye projeksiyonlar, Brain, 1967, 90, 521-540.
Macadar A. W., Rausch LJ, Wenzel B. M. Hutchison LV Güvercindeki koku alma yolunun elektrofizyolojisi, J. comp. Physiol., 1980, 137, 39-46.
McCabe BJ, Horn G., McGrath G. Civciv telensefalonunda asetilkolinesterazın dağılımı, J. Anat., 1982, 134, 600-601.
McCabe BJ, Hunt SP Peptitler ve evcil civcivin telensefalonunda biyojenik aminler. Bir immünosito-kimyasal çalışma, Neurosci. Lett., 1983, Ek. 14, S242.
Macphail EM Güvercinlerde intrakranial stimülasyondan pozitif ve negatif pekiştirme, Nature, Londra., 1967, 213, 947-948.
Meibach RC, Siegel A. Sıçandaki septal alanın efferent bağlantıları: retrograd ve anterograd taşıma yöntemlerini kullanan bir analiz, Brain Res., 1977, 119, 1-20.
Milner P. Hipokampus ve öğrenme - teori. İçinde: Bireysel Edinilmiş Davranış Biçimleri ve Merkezi Mekanizmaları. Gagra konuşmaları, 1987, c. 8, s / ed. M. M. Khananashvili (baskıda).
Mitchell/. A., Hali G. Güvercinlerde paleostriatal lezyonlar ve enstrümantal öğrenme, Quart. J. exp. Psychol., 1984, 36B, 93-117.
Mogensen J. t Divae I. Güvercinlerdeki prefrontal “korteks”. Davranışsal kanıt, Beyin Davranışı. Evol., 1982, 21, 60-66.
Mora R., Roll ET, Burton MJ Hipotalamustaki nöronların yiyeceğin görülmesine verdiği tepkilerin öğrenilmesi sırasında modülasyon, Exp. Neurol., 1976, 53, 508-519.
Morenkov ED, Khun DK Karga Corvus corone, J. evol'un beyninin hiperstriatal bölgesindeki görsel stimülasyona nöronal tepkiler . biyokimya Physiol., 1977, 13, 51-55.
Mountcastle VB Çevremizdeki dünya: seçici dikkat için nöral komut işlevleri, Neurosci. Res. Program Bull., 1976, 14, Ek.
Mountjoy CQ, Rossor MH, iversen LL, Roth M. Kortikal kolinerjik ve GABA eksikliklerinin senil demansta kantitatif nöropatolojik bulgu ile korelasyonu, Brain, 1984, 107, 507-518.
Nauta WJH Frontal lob sorunu: yeniden yorumlama, J. Psychiat. Res., 1971, 8, 167-187.
Parker DM, Delius JD Güvercinin ön beyin bölgesindeki görsel uyarılmış potansiyeller, Exp. Brain Res., 1972, 14, 198-209.
Pearson R. Kuş beyni, 1972. Academic Press, Londra.
Phillips RE Yeşilbaş ördeğindeki "Vahşilik": beyin lezyonlarının ve stimülasyonun "kaçış davranışı" üzerindeki etkileri, J. comp. Neurol., 1964, 122, 139-155.
Phillips RE, Youngren O. M. Beyin stimülasyonu ve türe özgü davranış: tavuk beyinlerinin (Gallus gallus) elektriksel stimülasyonuyla uyandırılan aktiviteler, Anim. Behav., 1971, 19, 757-779.
Raisman G. Hipotalamusun sinirsel bağlantıları, Br. tıp Bull., 1966, 22, 197-201.
Reiner A., Brauth SE, Karten HJ Bazal ganglionların evrimi, Trends Neurosci., 1984, 7, 320-325
Rieke GK Güvercin paleostriatumunun Kainik asit lezyonları: hareket bozukluklarının incelenmesi için bir model, Physiol. Behav., 1980, 24, 683-687.
Rieke G. K., Wenzel B. M. Güvercin koku ampulünün ön beyin projeksiyonu, J. Morph., 1978, 158, 41-56.
Ritchie T., Cohen DH Kuş tektofugal görsel yolu: telensefalik hedefinin projeksiyonu, ektostriatal kompleks, Proc. sos. Neurosci., 1977, 3, 94.
Russell J. R. DeMeyer W. Macaca rhesus'un piramidal aksonlarının kantitatif kortikal kökeni , Neurology, 1961, 11, 96-108.
Sahgal A. Hipokampal lezyonlar, güvercinlerde tanıma hafızasını bozar, Behav. Brain Res., 1984, 11, 47-58.
Scheich H. Civcivlerin işitsel neostriatumundaki iki sütunlu sistem: 2-deoksiglikozdan kanıtlar, Exp. Brain Res., 1983, 51, 199-205.
Scheich H., Bonke BA, Langner G, Gine tavuğundaki bazı işitsel çekirdeklerin 2-deoksiglukoz tekniği Celi ile gösterilen fonksiyonel organizasyonu. Doku Res., 1979a, 204, 17-27.
Scheich H., Langner G., Bonke D. Gine tavuğunun (Numida meleagris) işitsel neo striatumundaki birimin türe özgü çağrılara ve sentetik uyaranlara duyarlılığı. II: iambus benzeri aramaların ayrımı, J. comp. Physiol., 1979b, A132, 257-276.
Scheich H., Bock W., Bonke D., Langner G., Maier V. Gine tavuğunda (Numida meleagris) akustik iletişim . In: Advances in neuroethology (editörler J.-P. Ewert, RR Capranica ve DJ Ingle), 1983, s. 731-782, Plenum Press, New York.
Shiosaka S., Takatsuki K., Inagaki S., Sakanaka M., Takagi H „ Senba E., Matsuzaki T., Tohyama M. Topografik atlas of somatostatin-containing neuron system in the avianbeyin ve ilişkili olarak katekolamin içeren nöron sistemi . II. Mesencephalon, eşkenar dörtgen ve omurilik, J. comp. Nörol.,
202, 115-124.
Squire LR, Cohen NJ, Nadel L. Medial temporal bölge ve bellek konsolidasyonu: yeni bir hipotez. İçinde: Bellek birleştirme (editörler M. Weingartner ve E. Parker), 1984, s. 185-210, Erlbaum, Hillsdale, NJ
Vinogradova OS Limbik sistemin Bilgi kaydındaki rolü. gerçekler ve hipotezler. In: The Hippocampus (editörler K. Pribram, R. Isaacson), 1975, v. 2, s. 3-70, Bergama Basını, NY
VinogradovaOS Tartışması. Septohipokampal sistemin işlevlerinde, Ciba Vakfı Sempozyumu, 1978, Cilt. 58, s. 343-349. Alıntı Medica, Amsterdam.
Vowles DM, Beazley LD Kuşlarda duygusal davranışın nöral substratı. İçinde: Kuşlar, beyin ve davranış (ed. IJ Goodman ve M.W. Schein), 1974, s. 221-58. Akademik Basın, New York.
Wallenberg A. Der Ursprung des Tractus isthmo-striatus (öder bulbostriatus) der Taube, Neurol. Zentbl., 1903, 22, 98-101.
Warrington EK, Weiskrantz L. Amnezi: bir kopukluk sendromu mu? Nöropsikoloji, 1982, 20, 233-248.
Weiskrantz L. Hafıza kaybı çalışmalarının karşılaştırmalı yönleri, Phil. Trans. R. Soc.,
B298, 97-109.
Whitehouse P. L, Price DL, Struble RG, Clark AW, Coyle JT, DeLong MR Aİzheimer hastalığı ve senil demans: bazal ön beyinde nöron kaybı, Science, 1982, 215, 1237-1239.
Wilson P. Evcil civcivde görsel hiperstriatumun organizasyonu. • Görsel projeksiyonun topolojisi ve topografyası, Brain Res., 1980, 188, 319-332.
Wurfz RH, Goldberg ME Uyanık maymunlarda göz hareketleriyle ilgili üstün kollikulus hücre yanıtları. Bilim, 1971, 171, 82-84.
ve RH t Goldberg ME, Robinson DL Mönkey'de görsel yanıtın davranışsal modülasyonu: dikkat ve hareket için uyaran seçimi, Progr.
7 Psikobiyol. fizik. Psychol., 1980, 9, 44-83.
4e,er H., Karten HJ Güvercin arşivi: afferent ve efferent bağlantıların organizasyonu, Brain Res., 1971, 31, 313-326.
BÖLÜM 12
SENTEZ YOLUNDA
Önceki bölümlerin ayrıldığı çalışmayı özetlerken, iki soru sorulmalıdır. Bu çalışma, hafıza mekanizmalarının anlaşılmasına ne şekilde katkıda bulundu ve damgalamanın anlaşılmasına ne şekilde katkıda bulundu?
Hafıza
Nöropsikolojik ve nöroetolojik bakış açıları
geçmiş bir olayın izlerinin korunmasının "... yollardaki özel değişikliklerin, yani en iyi süreçlerin varlığında morfolojik tezahürlerle" ifade edildiği şeklindeki görüşüne atıfta bulunarak hafızanın analizine yeniden başlamakta fayda var . beyin dokusu" (James, 1890, cilt I, s. 655). Ancak James'in hafıza hakkında bundan çok daha fazlası vardı. Bir "görüntünün" güncel bir olaydan çok geçmiş bir olaya atıfta bulunduğunu kabul etmek için "... bu görüntüyü birbiriyle bağlantılı bir dizi çağrışımla ilişkilendirmenin" gerekli olduğuna işaret etti (James, 1890, cilt I, s.650) . Bu ifadeyi , n'nin geçmiş bir olayı, o'nun bir "bağlam", örneğin eşlik eden olayları ve r'nin geçmiş bir olayı hatırlamaya sevk eden bir gerçek olduğu bir diyagramla açıkladı. m, l ve o düşünceleri tarafından harekete geçirilen "sinir merkezleri" sırasıyla M, N ve O olarak etiketlenir (Şekil 12.1). Bir n olayı , M N ve O'yu heyecanlandırıyorsa meydana geldiği bağlamla birlikte hatırlanabilir . _
James modeli, beyin hasarının neden olduğu hafıza bozukluğunu analiz etmede yararlıdır, özellikle de model, onun beceri oluşumuyla ilgili bazı fikirlerini içerecek şekilde genişletilirse. James, bir beceriyi , "kullanımı" impulsların iletilmesini kolaylaştıran sinir yollarındaki deşarjlara dayanan, sık sık tekrarlanan bir eylem olarak tasarladı . Like Model (Maudsley, 1876), bir becerinin gerçekleştirilmesine inanıyordu.

Resim: 12.1. William James'in "... hafızanın tüm kökenini açıklayan" diyagramı. M, N ve O daireleri "sinir merkezleridir". N bazı geçmiş olayları temsil eder; O - gerçekleştiği bağlam; M, geçmiş olayın hatırlanmasına yol açan "gerçek" dir. (James, 1890, Cilt I, s. 655.)
beynin "yüksek" kısımlarının çok az veya hiç dahil edilmesini gerektirmez. Bazı ilişkisel öğrenme türleri, bir becerinin özelliklerine sahiptir; bu özellikler , uyaran tarafından uyarılan bölgeler arasındaki muhtemelen multisinaptik bağlantıların güçlendirilmesini ve tepkinin kontrol edilmesini açıklayabilir . Olası bir yolun bir örneği Şekil 1'de gösterilmektedir. 6.6, B,
belirli bir reaksiyonu kontrol eden hücreler arasındaki bağlantıların işlevsel verimliliğindeki bir değişiklik, bu uyaranın öğrenmeden önce mevcut olmayan belirli bir sinirsel temsilinin oluştuğu anlamına gelir. Bu anlamda sinir sisteminde uyaranın izi veya ilkel bir hafızası vardır. Bu bellek, yerel bir nöron zincirinde depolanır. Alışkanlığın bazı basit biçimleri de benzer şekilde yorumlanabilir. Alışkanlık, yeni bir uyarana yönlendirme refleksi veya bir vücut parçasını deri uyaranından çıkarmaya yanıt gibi öğrenilmemiş bir tepkinin, uyaranın tekrarlanmasıyla kademeli olarak zayıflaması sürecidir. Bu durumda, uyarıcı ve tepkiyi birbirine bağlayan yollar daha az verimli hale gelir (Bölüm 1.1). İlişkisel öğrenme ve alışma birçok yönden farklılık gösterir, ancak işlevsel olarak ortak bir noktaları vardır: öğrenme, uyaranın davranışı etkileme yeteneğinde bir değişikliğe yol açar. İlişkisel öğrenme , uyarıcı ve tepki arasındaki bağlantının artmasına, alışma ise zayıflamasına neden olur. Bazı çağrışımsal öğrenme ve alışma biçimleri , sinir sisteminin yalnızca bir kısmı sağlam kaldığında bile ortaya çıkabilir (Horn, 1970). Asgari gereksinimler açık olsa da, sinir sisteminin tam olarak hangi bölümünün bozulmadan kalması gerektiği açık değildir: uyaranları ayırt etmek için yeterli duyusal aparat, bir yanıtı düzenlemek ve yürütmek için yeterli motor sistem ve bazı çağrışımsal öğrenme biçimleri söz konusu olduğunda, motivasyon sisteminin yeterli bir parçası (Bölüm 6.4, 9.1 ve 9.3).
PMVG'de iki taraflı lezyonları olan civcivlerin yanı sıra medial temporal lob veya diensefalon hasarı olan insanlar ve maymunlarda basit beceriler öğrenilebilir . Primatlar söz konusu olduğunda, insanlarda ve maymunlarda görülen amneziler arasındaki benzerlikleri bulmak için çok çaba sarf edilmiştir (örneğin bkz. Weiskrantz, 1966-1982; iversen, 1976, 1983). Bir yaklaşım, 1) becerileri gerçekleştirmek için gereken bellek ve 2) bu becerilerin sonuçlarını depolamak için gereken bellek ( Ryle, 1949) ile nesneleri tanırken olduğu gibi beceri dışı tip bilgileri birbirinden ayırmak olmuştur. İlk bellek biçimi "prosedürel", ikincisi - "bildirimsel " olarak adlandırılır (Cohen ve Squire, 1980; Zoia-Morgan ve Squire, 1984). Amnezilerde "işlemsel" bellek esas olarak korunurken, "bildirimsel" bellek bozulur . "İşlemsel" bellek terimi, tüm eksikliklerine rağmen (Weiskrantz, 1986), hem insan hem de hayvan çalışmalarında uygulanmasını kolaylaştırır; Deneyimi sözlü olarak ifade etme yeteneğini ifade eden "bildirimsel" bellek terimi, böyle bir aktarıma izin vermez . Daha basit "dönüşlü" ve "yansıtıcı" bellek terimlerinin (L. Weis krantz, kişisel iletişim) , ayrışması medial temporal lob hasar gördüğünde meydana gelen iki tür bellek arasındaki temel farklılıkları daha etkili bir şekilde yansıtması mümkündür. "yansıtıcı" bellek terimi, hayvan öğrenmesi üzerine çalışanlar tarafından evrensel olarak kabul edilmeyebilecek ek nüanslara sahip olsa da .
Amnezinin klinik belirtilerini inceleyen Warrington ve Weiskrantz (Warrington, Weiskrantz, 1982) , bunun iki bellek sisteminin ayrılmasının bir sonucu olduğunu öne sürdüler. Bunlardan birinde aracı bellek sistemi, “... izler değişebilir, birbirine bağlanabilir ve olayların sürekli değişen kaydı şeklinde saklanabilir. Normal kişinin geçmiş deneyimlerle ilgili olayları hatırlaması ve tanıması , bu aracı bellek aracılığıyla mümkün olabilir . Bağlamsal bağlantılar açısından zengin izler depolayan ve insanlarda amnestik sendrom sırasında zarar gören bu sistem, N sistemlerinin bazı özelliklerini taşır . ve James tarafından öne sürülen O (Şekil 12.1). İkinci bellek sistemi, geçmiş deneyimlerin nispeten değişmez izlerini saklar ve böylece dış dünya hakkında istikrarlı bilginin temelini oluşturur. Bu sistem anlamsal bellek olarak adlandırılmıştır (Tulving, 1972). İnsanlarda amnezik sendromdan etkilenmez. Dış dünya hakkında bazı bilgilerin ilişkisel öğrenme yoluyla edinilmesi mümkündür ( örneğin, "Kırmızı ışıkta dur"). Bu tür bir hafızanın sinirsel temeli, alışkanlıkların ve alışkanlıkların oluşumunda yer alan hafızanın temelinden belki de çok az farklıdır. Ancak açık bir şekilde kelimelere veya sembollere atıfta bulunan "semantik" terimi , insan hafızasının bazı yönlerini tanımlamada geçerli olmakla birlikte , hayvanlarda hafızayı tarif etmek için uygun değildir. İnsan ve hayvan hafızası çalışmalarına eşit derecede uygulanabilir bir terim bulunana kadar, belki de bu ikinci sistem basitçe "dolayımsız hafıza" olarak adlandırılmalıdır. Aracı bellek, gerçekleştirdiği işlemlerin doğasına (Warrington ve Weiskrantz, 1982, s. 242) ve belirli beyin lezyonlarına, özellikle insanlarda ve maymunlarda medial temporal lob hasarına karşı duyarlılığı temelinde tanımlanır ; "aracısız bellek" bu tür hasarlara karşı bağışıktır.
Yukarıda bahsedildiği gibi, becerilerin uygulanmasında yer alan bu "aracısız hafıza", uyaran ve tepkiyi birbirine bağlayan sinir bağlantılarının güçlendirilmesine dayanabilir. Ancak mevcut bir beceri temelinde yeni beceriler oluşturulabilir. Aynı zamanda, yeni öğrenme durumlarında nasıl kullanıldığını açıklamak için bir becerinin uygulanmasının sonucunun içsel bir temsilinin oluşumuna izin vermek gerekir (bkz. Dickinson, 1980; Mackintosh, 1985). Her iki bellek biçimi de alışkanlık sırasında işlev görmelidir. Örneğin, bir insanda cilt yüzeyinin hafifçe uyarılmasıyla oluşan karın kaslarının kasılmasına yönelik alışma refleksini ele alalım. Bu refleks , uyaranın tekrar tekrar sunulması üzerine giderek zayıflar (Hagbarth ve Kugelberg, 1958). Artık yanıt verme ihtiyacının kalmadığı anı, deri uyarımına yanıt veren duyusal nöronlar ile yanıttan sorumlu kaslar arasındaki iletimin bloke edilmesiyle temsil edilebilir (Bölüm 1.1). Bu tür bellek "aracısız" olarak adlandırılabilir. Bununla birlikte denek , uyaranın birden fazla kez uygulandığını ve karın kaslarının kasılmasına neden olmayı yavaş yavaş bıraktığını hatırlayabilir. Bu tür bir bellek şüphesiz "aracılı" olarak kabul edilebilir. Alışkanlık prosedürlerinin sadece insanlarda değil hayvanlarda da iki nöral temsil veya hafıza izi oluşturduğuna dair kanıtlar daha önce tartışılmıştı (Bölüm 1.1.5; ayrıca bkz. Mackintosh, 1986).
Makaklarda forniks kesildiğinde bazı becerilerin gelişimi bozulmazken bazılarının gelişimi bundan etkilenir (Gaffan ve ark., 1984). Forniks hasarından sonra devam eden beceri biçimleri nispeten basittir. Çalışmanın bir biçimi, A nesnesine takviye kuvvetleri eşlik ederken, B nesnesine eşlik etmemekti. Sonra birlikte sunuldular. Bir deneyde maymun, yarım fıstık almak için önceden güçlendirilmiş bir nesneyi (A) hareket ettirmek zorunda kaldı. Forniks hasarlı maymunlar, kontrol grubu kadar iyi performans gösterdi. Bununla birlikte, tepkilerin hayvanın "doğal eğilimlerine" zıt olduğu alışkanlıklar ihlal edildi (s. 144-145). Örneğin, bir maymun yukarıda açıklananla aynı şekilde eğitildi, ancak daha önce güçlendirilmemiş bir nesne B'yi hareket ettirmek gerekliydi. Bu görevin başarılı bir şekilde tamamlanması, yeniden düzenlemeyi , tepkilerde değişiklikleri ve dolayısıyla sözde "aracılı" belleğe erişimi gerektirebilir. sistem. Fornix zarar görürse bu erişim bozulur.
Warrington ve Weiskrantz ( 1982) , "aracılı" belleğin bazı kritik bileşenlerinin ön loblarda yer alan yapılar tarafından gerçekleştirildiğini ileri sürerler . Bu yazarlar, amnestik sendromun, bu bellek sistemini semantik ("aracısız ") bellek sistemine bağlayan bağlantılar koptuğunda ortaya çıkabileceğine inanmaktadır. Başka bir deyişle, bu tür bir hasardan sonra bazı beceriler edinilebilirse, bunların uygulanmasının sonucu "aracılı" bellekte sabitlenemez. Warrington ve Weiskrantz'ın fikirleri doğruysa, o zaman frontal korteksteki hasar, bozulmuş hafıza ile ilişkilendirilmelidir. Gerçekten de frontal bölgedeki bazı bağlantıların hasar görmesinden sonra amnezik bir sendromun ortaya çıkışı anlatılmıştır (Mabille ve Pitres, 1913). Hafıza kusurları sıklıkla frontal lob tümörlü hastalarda bulunur (Nossen, 1964). Bununla birlikte, bu bozuklukların özellikle frontal bölgelerdeki hasarla mı ilişkili olduğu yoksa örneğin artan kafa içi basıncın bir sonucu olarak beyin fonksiyonlarının daha genel bozukluklarından mı kaynaklandığı tam olarak açık değildir (Walsh, 1978; De Renzi, 1982). ).
Diğer bazı gözlemler bu çelişkilerin nedenini açıklayabilir. Aggleton ve Mishkin (1983b) , diensefalon hasarından sonra makaklarda hafıza bozukluğunu incelediler . Bir durumda talamusun dorso-medial çekirdeği, diğerinde anteroventral çekirdek hasar gördü. Her iki lezyon da nesne tanımada orta derecede hafıza bozukluğuna neden oldu. Her iki çekirdekteki birleşik hasar, tam amneziye yol açtı (Aggleton ve Mishkin, 1983a). Primatlarda, prefrontal korteks talamusun dorsomedial çekirdeği için ana hedeftir ve cingular korteks anteroventral çekirdek için ana hedeftir (Bölüm 11.3). Bu kortikal alanların her birinin hasar görmesi amneziye neden oluyorsa, maksimum etki bunların birleşik hasarıyla ortaya çıkar. Bu bağlamda, Whitty ve Lishman'ın (Whitty ve Lishman, 1966) raporu ilgi çekicidir.İnsanlarda beyin hasarının hafıza üzerindeki etkisine ilişkin incelemelerinde, hafıza kaybının, cingular korteksin ön kısmı bozulduğunda meydana geldiğini not ederler. kaldırıldı. Bu rahatsızlıklar, tam amnezidekinden daha az kalıcı ve daha az kesindi; Egleton ve Mishkin'in gözlemlerinden çıkması gereken sonuç budur .
, talamusun medial çekirdeklerinden yolların gittiği temporal lobun medial bölgesinin kortikal projeksiyonlarına bağlı olduğu varsayılabilir . Önemli olan, yalnızca primatlarda tamamen ayrılmış olan cingular ve prefrontal bölgelerdir (Şekil 11.12). Örneğin sıçanda, talamusun bu iki çekirdeğinin kortikal çıkıntıları yakından örtüşür (Divac ve diğerleri, 1978). Bu veriler , söz konusu bölgelerin işlevsel olarak örtüştüğünü önermemizi sağlar . Primatlarda , bu bölgeler ayrı olmakla birlikte birbirine sıkı sıkıya bağlıdır.
saniyede Şekil 11.3'te, bir yandan tavuklardaki PMCG bağlantıları ile diğer yandan primatlardaki prefrontal ve cingular korteks arasındaki bağlantılar arasındaki benzerliklere baktık. Kuşlar ve primatlar kadar farklı olan hayvanlardaki yapı ve işlevleri karşılaştırmak kesinlikle garip görünüyor . Bununla birlikte, beynin nesneleri tanıma ve primatlarda görsel farklılaşmalar geliştirme işlevlerine ilişkin deneysel çalışmalar ancak yakın zamanda geliştirilmiştir. Bu, Aggleton ve Mishkin (1983a ) tarafından makalelerinde bildirilen sonuçlara ilişkin bir tartışmanın ilk paragrafında belirtilmiştir: diensefalik amnezinin merkezi semptomları hakkında. Talamusun bu bölümü, korteksin prefrontal ve cingular bölgelerine uzanır. Ve bu bölgelere bazı anatomik benzerlikler taşıyan SMVG'deki hasar, bazı ilişkisel öğrenme biçimlerini etkilemeden görsel nesnelerin tanınmasını da bozar (Bölüm 6).
Mevcut veriler, sol SHG'nin kendisinin bir bilgi depolama yeri olabileceğini göstermektedir. Eğitimden sonra içinde bulunan sinapsların yapısındaki değişiklikler Bölüm 1'de açıklanmıştır. on; bu nedenle, onları burada tartışmayacağız. Bu veriler, bellek işlemlerinin PMIG dışında başka bölgelerde de yer alma olasılığını dışlamaz (aşağıya bakın), ancak bazı açılardan tanıma için gerekli izlerin depolanmasının beynin yerel bir alanı tarafından sağlandığını gösterir. . Görünüşe göre PMMG, primat beyninin aracılı hafıza sağlayan sistemin parçası olan bölgelerine karşılık geliyor . SMVG'nin ve muhtemelen bir bütün olarak tüm ventral hiperstriatumun daha fazla incelenmesi, hafıza izlerinin nasıl oluştuğuna ve depolanan bilgilerin aracılı hafıza sisteminde nasıl işlendiğine ışık tutabilir.
Tavuklarda sol LMCG, tanıma için gereken hafızada yer alan tek yapı değildir. X' (bölüm 7 ve bölüm 8.5) olarak adlandırılan başka bir sistemin söz konusu olduğuna şüphe yoktur. Bu iki sistemin bellekte aynı işlevlere sahip olduğunu varsaymak mantıksal olarak zordur; ancak, bu farkın nelerden oluştuğunu gösteren hiçbir deneysel veri henüz yoktur. İnsanlarda ve maymunlarda medial temporal lob hasar gördüğünde , hem tanıma için gerekli hafıza hem de bazı davranışsal tepkilerin sonuçlarını saklamak için gerekli hafıza bozulur (Zola-Morgan ve Squire, 1984). Bununla birlikte, bu iki bellek türü, bilgileri işlemek için tamamen farklı yollar gerektirebilir. Nesne tanıma bile tek bir işlem olmayabilir. Nesne bilgisi, az veya çok sayıda ilişkili çağrışımla birlikte saklanabilir (James , 1890 ; ayrıca bkz . Aracılı bellek biçimlerinin çeşitliliği hakkında doğrudan deneysel veri yoktur, belki de farklı bellek türleri hakkındaki nöropsikolojik fikirlerin yakın zamanda ortaya çıkması nedeniyle. Görünüşe göre, sol PMVG ve X' ile ilgili daha ileri çalışmalarda nöropsikolojik ve nöroetolojik yaklaşımları birleştirerek, bu fikirleri rafine edebileceğiz.
Nörofizyolojik yönler
Bir olay hakkında bilgi saklarken, bir şekilde bir zaman damgası alıyor gibi görünüyor. Buna duyulan ihtiyaç, bir olayın meydana geldiği bağlamın zamanı içerdiğine inanan James (James, 1890) tarafından sezgisel olarak anlaşıldı . Amnezi olan hastalar , deneyimlerinin zaman içindeki sıralamasına bağlı olan görevlerin yerine getirilmesinde ciddi şekilde bozulmuştur (Huppert ve Piercy, 1978). Zaman damgaları şüphesiz diğer birçok hayvanın hafızasında mevcuttur. Örneğin, bir hayvan nektar gibi yenilenebilir bir besin kaynağı kullanıyorsa (bkz. örneğin Kamil, 1978), yalnızca kaynağın ziyaretinin değil, o ziyaretin zamanının da takip edilmesi önemlidir. Bu nedenle, ikinci ziyaret , besin kaynağının restorasyonu ile aynı zamana denk gelecek şekilde zamanlanabilir . Zaman duygusunun fizyolojik temeli ne olabilir ? Sinir sisteminde saat benzeri özelliklere sahip nöronlar var mı?
Strumwasser ( 1973), izole edilmiş bir Aplysia parietovisceral gangliondaki nöronların aktivitesini kaydetmiştir. İzole edilmiş bir ganglion kültürde birkaç hafta muhafaza edilebilir. Birçok hücrede ritmik deşarjlar gözlendi. Ritimler yaklaşık 24 saatlik bir periyodikliğe sahipti , ancak bu periyotlar farklı hücrelerde değişiklik gösteriyordu (Jacklet, 1981). Tavşanların (Vinogradova, 1970) ve kedilerin (Nogi, 1962) merkezi sinir sisteminde zaman içinde oldukça doğru bir deşarj dağılımına sahip nöronlar da kaydedilmiştir . Örneğin, anestezi uygulanmamış ve kısıtlanmamış bir kedinin görsel korteksindeki nöronal aktiviteyi kaydederken , çok nadir fakat şaşırtıcı derecede düzenli deşarjlar veren küçük bir hücre grubu tanımlandı. Tespit edilen beş hücrenin her biri için darbeler arası aralıkların dağılımı, şekil 2'de gösterilmiştir. 12.2. Tüm bu hücreler aynı hayvan üzerinde kaydedilmiştir. Hücreler, birkaç saniyeye kadar süren bir duraklamanın ardından gelen tekli artışlarla boşaltıldı. Her hücre için, dikenler arası aralıklar çok dar bir dağılıma sahipti. Bazen iki aksiyon potansiyeli arasında çok uzun aralıklar vardı, ancak süre olarak her zaman belirli bir hücre için standart aralığın iki veya üç katına yakındı (Şekil 12.2, kutu 2, 3 ve 4). Görsel veya diğer duyusal uyaranlar deşarjların sıklığını etkilemedi. Bu nöronlar, onlarla aynı anda kaydedilen, ancak duyusal uyaranlardan birine yanıt veren diğerlerinden keskin bir şekilde farklıydı.
Düzenli deşarjlı hücrelerin etki mekanizması iki basit varsayım temelinde analiz edildi. Bir yandan, bu hücreler yankılanma aktivitesine sahip çok uzun bir nöron zincirinin parçası olabilir. Bununla birlikte , sırasıyla sağ ve sol hemisferlerde kayıtlı olan 2 ve 5 numaralı hücreler hariç (Şekil 12.2), her bir hücredeki aralıkların ortalama değeri diğerlerinden önemli ölçüde farklıydı (P<0.001). Kayıt tek bir hayvandan yapıldığından, her biri diğerlerinden bağımsız olarak çalışan yankılanma aktivitesine sahip en az dört benzer devre olduğunu varsaymalıyız . Öte yandan, deşarjların bu doğası , beş hücrenin her birinde işleyen bir tür sinaptik olmayan kimyasal süreci yansıtıyor olabilir. Gerçekten de Strumwasser ( 1973) , sinaptik etkiler farmakolojik maddeler tarafından bloke edildiğinde Aplysia nöronlarının ritmik deşarjlarının korunduğunu gösterdi .
140
Kletna 4, P. Hücre 5, L.
Ortalama = 9,42 Ortalama = -0,25
SD=0,72 SD=0,67
N=51 N=61
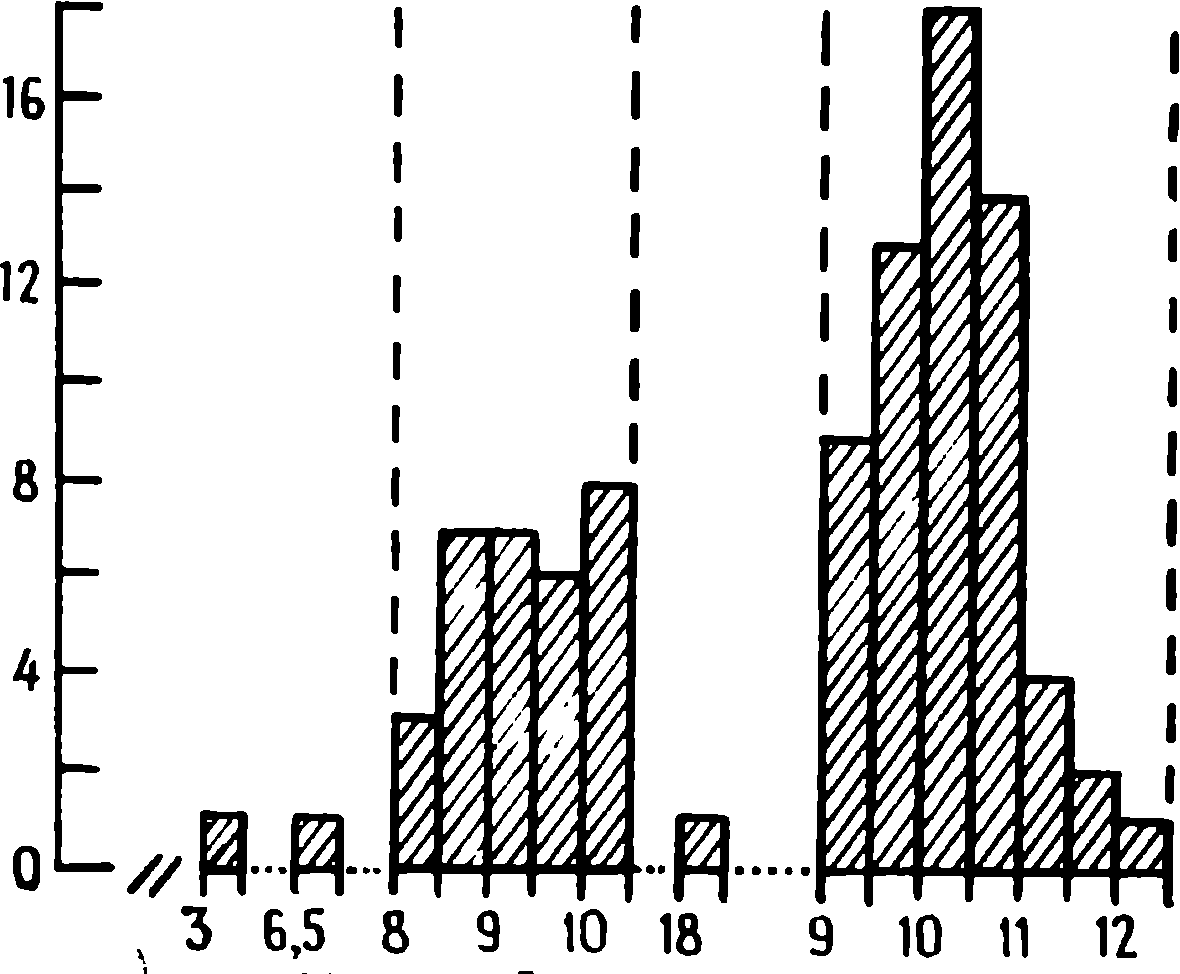
, eğer gerçekten önemliyseler, bir olayın sinirsel temsilini işaretlemek için bunların nasıl kullanılabileceğini açıklamaz . Beyinde bilgi depolamanın fizyolojik mekanizmalarını analiz etmeye çalışırken bu sorun şüphesiz dikkate alınmalıdır .
Bir olayın meydana gelme zamanı, izleriyle birlikte depolanan tek bağlamsal bilgi biçimi değildir. Hafızada diğer nesnelerin ve olayların varlığını veya yokluğunu, bunların uzamsal düzenini, sırasını vb. Sabitlemek de önemlidir. Merkezi sinir sisteminde tepkileri alınan bağlamsal bilgiye bağlı olabilen nöronlar varsa, o zaman her şeyden önce onlar tabii ki temporal lobun medial bölgesinin hipokampal bölgesinde aranmalıdır . Bu tür nöronları araştırmayı ve incelemeyi amaçlayan deneylerde, rhesus maymunlarına önce S1 uyaranı ve ardından S2 uyaranı sunuldu (Brown ve Homni, 1978; Brown, 1982). Uyarıcılar büyük veya küçük daireler ve karelerdi. Maymunlar, S1 ve S2 aynı şekil ve boyuttaysa sağ panele basmak üzere eğitildi ve doğru tepkiler meyve suyuyla pekiştirildi. S1 ve S2 şekil olarak eşleşiyor ancak boyut olarak farklıysa , maymun sol panele basmış olmalıdır. Örneğin, belirli bir denemede hem S1 hem de S2 büyük karelerse, doğru yanıt sağ panele basmaktı; ancak S1 küçük bir kare ve S2 büyük bir kare ise, o zaman doğru olan sol panele basmaktı (Şekil 12.3). Eğitim sırasında ve nöral aktivitenin kaydedilmesi sırasında, hayvan primatolojik bir sandalyedeydi ve panellere ve uyaranların göründüğü osiloskop ekranına dönük oturuyordu . Bazı hücrelerin belirli bir uyarana verdiği yanıtların S1 ve S2 olarak sunulmasına bağlı olarak farklılık gösterdiği bulunmuştur. Örneğin, S1'in büyük bir kare olması koşuluyla, S2 küçük bir kare olduğunda bir hücrenin deşarj sıklığı keskin bir şekilde arttı (Şekil 12.3d). S1 aynı zamanda küçük bir kareyse (Şekil 12.3, B) veya S1 ve S2 büyük karelerse (Şekil 12.3, K) bu artış önemli ölçüde daha azdı. Başka hiçbir uyaran kombinasyonu, aktivitede gözle görülür bir artışa neden olmadı (Şekil 12.3, C, E- G). Sonuç olarak, bu hücre için yanıt, yalnızca doğrudan sunulan uyaran (S2) veya maymunun hareket yönü (sağa veya sola) tarafından belirlenmedi, aynı zamanda küçük karenin göründüğü bağlama da bağlıydı.
tekrarlanan sunum üzerine tanıdık olarak tanınabilir . Rolls ve diğerleri (1982), 20* ile ilgili olabilecek hücreleri tanımlamıştır.


Resim: 12.3. Bir rhesus maymununun hipokampal bölgesinde kaydedilen bir hücrenin bağlama bağlı tepkileri. Doğru örneklerle ortalama aktivite histogramları verilir; dikey çizgiler standart hatanın değerini gösterir. İkinci uyaranın (S2) eylemi için sekiz farklı koşul gösterilir: büyük veya küçük resim, daire veya kare ; S2'den sonra sağ veya sol panele basma tepkisi . Beyaz ok - uyarı flaşı (süre 0,5 s). Taralı oklar, birinci uyaran S1 (1,0–1,5 s) ve ikinci uyaran S2'dir (2,0–2,5 s). Doğru test ile uyarı flaşı başladıktan 3-6 saniye sonra basılması yapılır. N, histogramda ortalaması alınan örnek sayısıdır. S2 uyaranının uyandırdığı nöral aktivite, sekiz koşul altında karşılaştırıldı. Farklılıklar anlamlıdır (P<0.001). Küçük kare uyaran olduğunda S2'ye yanıtta belirgin artışa ve ardından sol paneldeki (D) baskıya dikkat edin. Bu aktivite, S2'nin sağ panel (B) üzerindeki baskının eşlik ettiği küçük bir kare olduğu duruma göre (P<0.05) daha yüksekti ve S2'nin sağ veya sol panele baskının eşlik ettiği herhangi bir başka uyaran olduğu duruma göre önemli ölçüde daha yüksekti (P<0.01) . . Sonuç olarak, hücrenin yanıtı yalnızca o anda uygulanan uyarana (S2 küçük bir karedir) ve yalnızca motor tepkinin doğasına (sol panele basarak) değil, aynı zamanda uyaranın sunulduğu bağlama da bağlıydı. . (Brown ve diğerleri, 1982.)
8 bu tanıma süreci. Anestezi uygulanmamış rhesus maymunlarının ön talamuslarında serbest davranış koşulları altında hücresel aktivite kaydedildi . Bu deneyde maymuna bir nesne iki kez gösterildi. İlk sunulduğunda yeniydi. Bu durumda maymun, hoş olmayan salin solüsyonundan kaçınmak için ağzının önünde tüpü yalamak zorunda kalmadı. Maymuna daha sonra başka görsel uyaranlar verildi. Nesne ikinci kez ortaya çıktıysa ve bu nedenle tanıdıksa, maymun meyve suyu elde etmek için pipeti yalamış olabilir. Uyaranlara yalnızca tanıdık olduklarında yanıt veren bir nöron popülasyonu bulundu. Ek olarak, başka bir görevde - görsel farklılaşma - bu nöronlardan bazıları tanıdık bir uyarana hem olumlu hem de olumsuz pekiştirme ile yanıt verdi. Bu, uyaranların koşullu refleks farklılaşmasının gelişimi sırasında bile , nöronların, alınan takviyenin doğasına bakılmaksızın, tanıdık olduklarında onlara yanıt verdiğini gösterir.
Tabii ki, hafıza süreçlerinde yer alan nöronların aktivitesine ilişkin fizyolojik çalışmalar, yolculuklarının sadece başında. Ancak nöronları çalışılan hayvanlara doğru görevler verilirse , mikroelektrot kaydı bu sorunu uzun süre analiz etmek için değerli bir yöntem olarak kalacaktır .
baskı
Baskı oluşumunda ve görsel korteksin gelişiminde hassas dönemler
Bir civcivin hareketli bir nesneyi tercih etmesini sağlayan prosedürler ile yavru kedinin görsel korteksinin gelişimini etkileme yöntemleri arasında bariz bir benzerlik vardır (Horn ve diğerleri, 1973; Bischoff, 1983). Her iki durumda da, genç hayvana hassas bir süre boyunca bir uyaran sunulur ve ardından sinir sistemindeki ve davranışlarındaki değişiklikler kaydedilir. Bu prosedürler benzer olduğundan ve etkili etkilerinin süresi benzer olduğundan, şu soru ortaya çıkıyor: sonuçları arasında bir benzerlik olmayacak mı?
İlk olarak, hassas bir dönemde bir uyaranın nöronlar üzerindeki etkisini düşünün. Yavru kedilerde, yaşamın ilk haftalarında bir gözün kapatılması, bu gözün görsel korteks hücrelerinden işlevsel olarak ayrılmasına yol açar (Wiesel ve Hubel, 1963; Hubel ve Wiesel, 1970). Bu yetişkin kedilerde görülmez. Bu çalışmalardan çıkan sonuç , yavru kedinin görsel korteksindeki nöronların bir süre yüksek plastisiteye sahip olduğu ve plastisitenin yaşamın ilk ayının sonuna doğru arttığıdır (Bölüm 9.1). Bu süre zarfında, bir yavru kedinin görsel korteksindeki nöral reaksiyonların parametrelerini, bazen onu etkileyen görsel sinyalleri değiştirerek çok önemli ölçüde değiştirmek mümkündür. Hirsh ve Spinelli ( 1970), görsel ipuçlarını belirli bir yönelimdeki çizgilerle sınırlayan kedi yavruları için özel olarak tasarlanmış gözlükler kullandı. Bu yazarlar, bir göze görsel uyaran yalnızca dikey çizgilerden oluştuğunda, görsel korteksteki hücrelerin yalnızca o göze dikey çizgilerle uyarıldığında uyarıldığını bulmuşlardır. Bu göze sunulan yatay çizgiler görsel korteksin nöronlarında reaksiyonlara neden olmadı, ancak normal koşullarda yetiştirilen bir yavru kedide bu uyaran birçok kortikal hücreyi etkiliyor (Hubel ve Wiesel, 1962). Görsel korteks hücrelerinin tercih edilen oryantasyonel seçiciliği, uzunlamasına şeritlerden oluşan sınırlı bir görsel ortamda büyüyen kedi yavrularında da dikey çizgiler lehine kaydırılmıştır (Blakemore ve Cooper, 1970).
Kuş beyninin görsel Wulst'unun memelilerin görsel korteksine karşılık geldiği öne sürülmüştür (Nauta ve Karten, 1970). Bu bakış açısı, peçeli baykuşun görsel Wulst'undaki nöronal reaksiyonların özellikleri üzerine yapılan çalışmada bir miktar onay aldı (Pettigrew ve Konishi, 1976a). Bu bölgedeki nöronlar, birçok açıdan yetişkin kedininkine benzeyen alıcı alanlara sahiptir; tıpkı bir kedi gibi, her iki gözün de görsel olarak uyarılmasıyla çok sayıda nöron etkinleştirilebilir. Bu tür dürbünle etkinleştirilen nöronlar, bir baykuşun görsel Wulst'unda yoktur, tıpkı yaşamın erken dönemlerinde bir gözün dışlanmasından sonra yavru kedilerin görsel korteksinde de bulunmadığı gibi (Pettigrew ve Konishi, 1976b).
Tavuklardaki görsel Wulst nöronlarının alıcı alanları, görsel deneyim sınırlı olduğunda bir yavru kedinin görsel korteksinde olana benzer şekilde, damgalayıcı bir uyarana maruz kaldıklarında değişiyor mu? Ne yazık ki, bu sorunun net bir cevabı yok. Wilson (1980) , 5-6 haftalık tavuklarda görsel Wulst nöronlarının alıcı alanlarının özelliklerini tanımlamıştır . Milne, McCabe ve Horne (yayınlanmamış gözlemler) bu bölgede retinanın dağınık aydınlatmasına yanıt veren hücreler saptadı (ayrıca bkz . Payne ve diğerleri, 1984). Bununla birlikte, yerel alıcı alanlara sahip hücreler, damgalanmaya neden olan yapay bir nesneyle sunulan tavuklarda bile yaşamın 3. haftasından önce nadiren bulundu . Baskılamanın Wulst nöronlarının alıcı alanlarının özellikleri üzerindeki belirgin etkisinin olmaması dikkatle ele alınmalıdır. Bu çalışmalarda kullanılan anestezik ajanların genç tavuklarda Wulst nöronlarının tepkilerini baskılaması , ancak yaşlı kuşlarda etkilememesi olasıdır. Bununla birlikte, iki günlük civcivlerin optik çatısında lokal alıcı alanlara sahip hücreler bulunmuştur, bu nedenle anestezi gerçekten de nöronal reaktiviteyi bastırıyorsa, etkisi seçicidir.
PMWG'de, öğrenme sırasında nöronların yapısı değişir (Bölüm 10.2); Kayıt anestezi uygulanmış bir hayvanda yapılmış olsa bile bu alandaki hücrelerin spontan aktivitesi de değişir (Bölüm 10.1). PMWG'de yerel alıcı alanlara sahip hücrelerin aranması henüz önemli bir başarı ile sonuçlanmamıştır (s. 266-267). Bu nedenle, görsel olarak sınırlı bir ortamda yetiştirilen yavru kedilerin görsel korteksinde gözlemlenen alıcı alanların organizasyonundaki dramatik değişikliklerin , tavukların damgalama sırasındaki görsel Wulst veya PMVG'sinde bir analojiye sahip olduğuna dair henüz verimiz yok.
Kendisine belli bir nesne sunularak öğretilen bir tavuk, onu diğerlerine tercih eder; bu, sosyal davranışının değiştiği anlamına gelir. Bununla birlikte, yavru kedilerin, örneğin dikey çizgilerden oluşan görsel olarak sınırlı bir ortama maruz kaldıktan sonra bu ortama bağlılık gösterdiğine dair hiçbir kanıtımız yok, elimizde yok. Civciv yeni nesnelerden başarıyla kaçındığından ve karmaşık görsel uyaranları ayırt edebildiğinden (Bölüm 6.1) , damgalanmış civcivin genel görsel yetenekleri etkilenmiş gibi görünmüyor . Yavru kedilere gelince , ilk başta, yetiştirildikleri yöne dik olan konturlara göre genel olarak kör kabul edildiler (Blakemore ve Cooper, 1970). Daha yeni çalışmalarda , görsel davranışın bazı yönlerinin bozulmuş olduğu bulunmasına rağmen (Blasdel ve diğerleri, 1977; Kratz ve Lehmkuhle, 1983) , bu tür ciddi görme bozukluğu doğrulanmamıştır (Freeman ve Marg, 1975). Bu nedenle görsel bir uyaranın sunumu, civcivlerin ve yavru kedilerin davranışlarını farklı şekilde etkiler.
Monoküler yoksunluğun yavru kedilerin görsel korteks nöronları üzerindeki etkisi, daha önce kapalı olan gözün açılıp açık olanın kapatılmasıyla ortadan kaldırılabilir. Bu işlem hassas dönem bittikten sonra yapılırsa gözün baskınlığını değiştirmez (Blaketore ve van Sluyters, 1974). Bu sonuç, civcivlerin damgalama için hassas dönem tamamlandıktan sonra bir nesnenin sunumu üzerine öğrenme yeteneğinin korunmasıyla çelişir (aşağıya bakın).
, tavuklarda damgalamanın altında yatan belirli beyin yapılarının esnekliğinin , yavru kedilerin görsel korteksinde bulunan esnekliğe benzeme olasılığını dışlamaz . Bununla birlikte, tüm plastisite biçimlerinin sinir sisteminin işleyişinde benzer işlevlere sahip olduğuna inanmak için hiçbir neden yoktur. Bu, merkezi sinir sisteminin gelişimini analiz ederken netleşir. Embriyogenez sırasında maksimum plastisite sergiler, ancak plastik değişimlerinin derecesinin embriyodaki öğrenme ölçeğini yansıttığına inanmak saflıktan daha fazlası olacaktır. Öğrenme ve hafıza nöroplastisitenin göstergesi olsa da, plastisitenin tüm biçimleri öğrenme ve hafızanın göstergesi değildir.
Civcivler damgalamadan sonra yeni nesnelerden kaçındıklarında, bu nesneler hakkındaki bilgileri hatırlama yeteneği azalır. Bu nedenle, birkaç yıl önce Sluckin ve Salzen'in (1961) işaret ettiği gibi, damgalama kendi etkisinin bir sonucu olarak durmalıdır. Bununla birlikte, tavuklarda yeni nesnelerden kaçınma tepkisi, bu nesnelere sürekli maruz kalmakla zayıflayabilir. Bu koşullar altında orijinal baskı dönüşüme uğrayabilir. Salzen ve Meyer (Salzen ve Meyer , 1968) civcivlere yumurtadan çıktıktan sonraki ilk üç gün boyunca mavi bir top verdi. Mavi top, civcivin bulunduğu kafese asıldı . Eşzamanlı seçim testinde, civcivler tanıdık mavi topu yeni yeşil olana tercih ettiler. Daha sonra mavi topun yeşil topla değiştirildiği kafese geri döndüler. Üç günlük maruziyetten sonra civcivler onu orijinal mavi topa tercih ettiler. Daha sonra tekrar 3 gün boyunca her iki topun da çıkarıldığı kafeslere yerleştirildiler. Bu süre sonunda test edildiğinde civcivler ikinci nesneyi tercih ettiler; bu nedenle, orijinal tercihte bir yeniden düzenleme vardı ve bu yeniden düzenleme sabitti. Tavuklar asıl nesneyi unuttular mı? Görünüşe göre hayır, çünkü orijinal tercih uygun koşullar altında geri yüklenebilir (Cherfas ve Scott, 1981). Bu sonuç ve ikinci nesne tercihinin birkaç gün devam etmesi, her iki nesnenin de en azından tüm deney süresi boyunca sabit kalan nöral izlerinin oluştuğunu göstermektedir. Dolayısıyla bir nesnenin tercihinin değişmesi o nesneyle ilgili hafıza izlerinin silinmesi anlamına gelmez. Bu deneyler, civcivin bir cisim gösterildiğinde onun hakkında bilgi yakalayabildiğini ve tanımayı sağlayan hafıza sisteminin hassas dönem bittikten sonra bilgi saklama yeteneğini kaybetmediğini göstermektedir. Belirli bir nesnenin izlerini saklayan nöral topluluklar plastisitesini kaybedebilir. Durum böyle olmasaydı, bu nesnenin bellekteki temsili kaybolur veya ihlal edilirdi. Bununla birlikte, bu gruplar tarafından bir miktar esneklik korunabilir, bunun sonucunda yeni alınan bilgiler onlarla bağlantılar kurabilir ve saklanan bilgileri dönüştürebilir.
Civcivler hassas dönemin ötesinde öğrenmeye devam ederse , o dönemin kendisi ne olacak? Sadece annenin sinirsel temsili ile yaklaşma tepkisini düzenleyen sistem arasında bir bağlantı kurulmasından mı bahsediyor? Bu bağın güçlenmesinin sonucu , civcivin artan enerjiyle anneyi hızla takip etme yeteneğidir. Aynı zamanda yeni nesnelerden de kaçınacaktır (Şekil 7.11, ///). Anneye yakın olan civciv, yavaş yavaş kendisi ve çevrenin diğer ayrıntılarını öğrenir, yırtıcı hayvanların ve kendi türünün saldırgan üyelerinin saldırılarına karşı güvende olur. Hinde ( 1962, 1982) tarafından başka bir avantajdan bahsedilmektedir : damgalama için eğitim ihtiyacı, yumurtadan yeni çıkmış civcivdeki tanıma mekanizmalarının olgunlaşmamış olmasından kaynaklanmaktadır. Hynd'e göre yetişkin hayvanlar, mesafe veya görüş açısından bağımsız olarak bir nesneye aynı şekilde tepki verebilir , ancak bu yetenek kişisel deneyime bağlıdır. Annesini sadece karmaşık uyaranların kişisel kombinasyonları olarak gören bir civciv , bu "algısal sabitliğe" henüz sahip olmayabileceğinden, deneyimden bağımsız herhangi bir tanıma mekanizması çok karmaşık olacaktır. Hassas bir döneme sahip olmak, civcivin algısal istikrarı hala gelişirken güvenli bir şekilde bağlanmasını sağlar. Daha sonra civciv dış dünyaya zaten aşina olduğunda anneye olan bağlılığı zayıflar. Fizyolojik düzeyde, bu süreç alışkanlığın, belki de sinaptik iletimin bir depresyonunun ve hatta yaklaşma sistemi ile anneyi temsil eden nesnenin sinirsel temsili arasındaki bağlantıdaki sinapsların gerilemesinin sonucu olabilir (Şekil 7.11, /) . /). Bu nesneyle -gerçek anne ya da yapay uyaranla- uzun süreli yakın temas, kademeli sinaptik değişiklikler yoluyla (Bölüm 10.2), civcivin sinir sistemindeki izlerin sabitlenmesini sağlayabilir. Ücretsiz depolama sisteminin ilk gelen bilgilere tepkisi, daha sonra girilen bilgilere göre daha yoğun olabileceğinden , bu izin özellikle güçlü olması da mümkündür . Bu şekilde, bir hayvanın erken yaşam deneyimleri , cinsel partner seçimi de dahil olmak üzere gelecekteki davranışları üzerinde biçimlendirici bir etkiye sahip olabilir (Lorenz, 1935, 1937; Bambridge, 1962; Guiton, 1962, 1966; Schutz, 1965; Salzen, 1966). ;Bateson, 1980) .
12.2.2.. Yatkınlık ve öğrenme
Birçok hayvan, bazı nesnelere diğerlerinden daha güçlü tepki verme veya daha güçlü tepki verme eğilimi gösterir ve civcivler bu açıdan bir istisna değildir (Seligman ve Hager, 1972; Hinde ve Stevenson-Hinde, 1973). Bu tür eğilimlerin nöral mekanizmaları çok az çalışılmıştır. Bir tavuğun kırmızı kutu veya doldurulmuş tavuk tercihi üzerindeki PMVG hasarının etkilerindeki fark, böyle bir çalışmanın başlangıç noktasıydı, ancak bu durumda bu çalışma tamamlanmaktan çok uzak.
Hassas bir eş zamanlı seçim testi kullanarak, karmaşık bir görsel örüntüye maruz kalan tavuklarda doldurulmuş tavuk tercihinin daha hızlı geliştiğini bulduk. Doldurulmuş tavuk tercihi, daha yavaş da olsa, birkaç saat dağınık ışığa maruz kaldıktan sonra veya mümkünse aynı süre karanlıkta serbestçe hareket ettikten sonra da gelişir. Böyle bir deneyimin yokluğunda, bir tercih yoktu. Bu gelişen tercihi sağlayan nöronal sistem, işlevsel hale gelmeden önce bazı etkilerin onu etkinleştirmesi gerekiyormuş gibi davranır. Uygun lokomotor aktivite veya basit görsel deneyim veya bu deneyim biçimlerinin her ikisi birlikte bu dahil etme sürecine katkıda bulunur (Bölüm 8.2.2). Ortaya çıkan bu tercih için PMVP'ye gerek yoktur; muhtemelen aktivitesi testosteron tarafından değiştirilebilen spesifik bir nöron sistemine bağlıdır (Bölüm 8.4.1 ve 9.6). Bu bağlamda, Lorentz'in ( 1935, 1937) damgalama konusundaki görüşleri özellikle ilgi çekicidir . Damgalamayı "... bir bütün olarak türlerinin üyelerine yönelik, ancak bireysel nesnelere değil " yanıtları yöneten "... çok tuhaf bir öğrenme süreci ..." olarak görüyor . Bu davranış, farazi spesifik bir sistem tarafından düzenlenebilir, ancak evcil tavuklarda bu spesifik sistemin bir tür tanıma sistemi olacak kadar ince ayarlı olmadığı vurgulanmalıdır (Bölüm 8.3). Belki de Lorenz'in damgalama tanımı aşırı derecede kısıtlayıcıdır, çünkü en azından evcil tavuklarda yatkınlık ve kişisel deneyimin etkisi arasında bir birlik vardır. Doğal koşullar altında civcivler, çevrelerinin karmaşık görsel özelliklerine bir kez alıştıktan sonra , kendi türlerinin belirli özellikleri veya özellikleri için hızla bir tercih geliştirirler. Daha sonra öğrenme süreci bu tercihi sınırlayabilir ve onu belirli bir bireye yönlendirebilir. Böylece, ilk tercih, civcivi büyük olasılıkla gerçek annesi olan uygun nesneye yönlendirecektir. Daha sonra öğrenme süreci başlayacak ve civcivin onu tanımasına izin verilecektir. Bazı kuşlarda şarkı oluşumu sırasında yatkınlık ve öğrenme etkileşimleri de gözlemlenmiştir (Maur, 1984) ve genel olarak bu tür etkileşimler çok yaygındır (Hinde, 1982).
Hangi sinir mekanizmaları varsayılan spesifik sistemin özelliklerine sahip olabilir? Son zamanlarda, maymun temporal korteksindeki bazı nöronların çok özel tepkilere sahip olduğu bulunmuştur (Bruce ve diğerleri, 1981; Perrett ve diğerleri, 1982). Perrett ve diğerleri (1982) , al yanaklı maymunların temporal kortekslerinin sınırlı bir bölümündeki hücrelerin yaklaşık %10'unun yüzlere oldukça duyarlı olduğunu bulmuşlardır . İnsanların ve maymunların gerçek yüzlerine veya görüntülerine verilen bu tepkiler, koyu çizgiler, basit geometrik şekiller veya karmaşık üç boyutlu nesnelerin verdiği tepkilerden 2 ila 10 kat daha güçlüydü. Bazı hücrelerde, yüz çevrildiğinde , rengi veya boyutu değiştiğinde reaksiyonların şiddeti neredeyse hiç değişmiyordu. Yüzün bölümlerini ayrı ayrı kapatarak veya göstererek, farklı hücrelerin farklı bölümlere veya gruplara tepki verdiğini göstermek mümkündü. Ayrıca , hücrelerin bir kısmı için, yüz özelliklerinin doğru görüntüsüne verilen tepkiler, bunların karıştırıldığı kontrol çizimlerinden daha güçlüydü. Perrette ve meslektaşları, bu nöronları yüzleri veya bireysel özelliklerini kodlamada uzmanlaşmış bir sistemin parçası olarak görüyorlar. Bununla birlikte, bu nöronların belirli bireylerin yüzlerini kodladığına dair bir kanıt yoktur. Görünüşe göre, bu tür nöronlar beyinde çok az . Nasıl oluştukları veya yeni doğan maymunların şakak lobunda bulunup bulunmadıkları hakkında hiçbir şey bilinmiyor. Bununla birlikte, bazı fizyolojik kriterlere göre, yeni doğan çocukların insan yüzünün doğru şematik temsilini "karma" bir görüntüye tercih etmeleri (Gören ve diğerleri, 1975) ve gerçek durağan bir yüzü şematikten daha yakından incelemeleri ilginçtir. temsil ( Stechler, 1965).
ortaya çıkmasından sorumlu olan PMGB dışındaki nöronların varsayımsal sisteminin , primatlardaki "yüz algılayıcı nöronlara" yakın özelliklere sahip olduğunu öne sürmek çok caziptir. Böyle bir sistemin bir civcivde yumurtadan çıktıktan hemen sonra işlevsel hale gelmesi olası değildir (Bölüm 8.2.2). Bununla birlikte, bu sistemdeki sinapsların, civciv aktif olarak hareket etmeye başlar başlamaz işlevsel hale gelmesi beklenmelidir ve bunların aktivasyonu muhtemelen testosteronun katılımıyla gerçekleştirilir (bölüm 10.3.7). Bu öznitelik çıkarma sisteminin nöronları, karmaşık bir görsel ortamın belirli öğeleri tarafından etkinleştirilebilir ve en etkili şekilde , optimuma yakın gerekli tetikleyici özelliklerin bir kombinasyonuna sahip olabilen gerçek anne tarafından uyarılabilir. Bu özelliklerin ne olduğu deneyle bulunacaktır, ancak bunların bir göz ve bir kafa kombinasyonundan veya belki de belli bir çerçeve içinde birleştirilmiş iki daire ve birkaç çizgiden fazlasını içermesi olası değildir. Bu sistemi çalıştırdıktan sonra , yaklaşım reaksiyonlarını hala deneysel olarak incelenmesi gereken şekillerde düzenleyebilir .
Daha fazla araştırma için bir çağrı muhtemelen tartışmamızı sonlandıracaktır. Bu çağrı, üstlenilen araştırmanın deneysel, keşfedici doğasını vurgulayarak, bu kitabın sayfalarından defalarca duyulmuştur . Beyni incelemeye başladığımızda şaşırtıcı sonuçlarla karşılaşırız. İnterhemisferik asimetriler, çoklu bellek depoları , çeşitli fizyolojik sınırlamalar ve diğer birçok bulgu, bize ele alınması gereken zorluklar sunar. Beyni inceleyerek, bilgileri kaydetmeyi, depolamayı ve geri almayı, örüntü tanımayı, yaklaşma ve kaçınma tepkilerinin uygulanmasını, dikkati ve motivasyonu mümkün kılan mekanizmalarına erişim kazanırız. Mühürlemenin meydana geldiğini gösteren şaşırtıcı ama görünüşte basit davranışların meydana gelmesiyle ilgili etkileşimlere ilişkin araştırma henüz emekleme aşamasındadır.
Özet
Önceki bölümlerde açıklanan çalışmaların sonuçları özetlenmiştir. İnsanlarda ve maymunlarda özellikle temporal lobun medial bölgesinde ve diensefalon yapılarında beyin hasarı durumunda hafıza bozuklukları düşünülür . Bu kusurlar , tavuklarda PMVG'ye verilen hasarın sonuçlarıyla ilişkilidir. Becerinin yürütülmesi için gereken bellek ve tanıma için gereken bellek farklıdır. Bağlamın önemi vurgulanır; yanıtları bağlama bağlı olan nöronların özellikleri ve tanıdık nesnelere yanıt veren nöronlar açıklanmaktadır. Yavru kedilerin görsel korteksindeki baskı ve plastisite karşılaştırılır. Yatkınlığın altında yatan olası nöral mekanizmalar, doğuştan gelen eğilimlerin ve öğrenme sürecinin nasıl etkileşime girdiği ve türleri ve bireysel farklılıkları tanımadaki rolleri ele alınır.
EDEBİYAT
Aggleton J. R., Mishkin M. Maymunlarda medial talamik lezyonları takiben görsel tanıma bozukluğu, Neuropsychologia, 1983a, 21, 189-197.
Aggleton JP, Mishkin M. Maymunlarda sınırlı medial talamik lezyonları takiben hafıza bozuklukları, Exp. Beyin Res. 1983b, 52, 199-209.
Bambridge R. Evcil tavukta erken deneyim ve cinsel davranış, Science, 1962, 136, 259-260.
Bateson P. Japon Bıldırcınlarında optimal çiftleşme ve cinsel tercihlerin gelişimi, Z. Tierpsychol., 1980, 53, 231-244.
Bischoff H.-J. Damgalama ve kortikol plastisitesi: karşılaştırmalı bir inceleme, Neurosci. Biyodavranış. Rev., 1983, 7, 213-225.
Blakemore C., Cooper GF Beynin gelişimi görsel ortama bağlıdır, Nature, Londra., 1970, 228, 477-478.
Blakemore C., van Sluyters R.C. Yavru kedilerde monoküler yoksunluğun fizyolojik etkilerinin tersine çevrilmesi: hassas bir dönem için daha fazla kanıt, J. Physiol. Londra., 1974, 237, 195-216.
Blasdel GG, Mitchell DE, Muir DW, Pettigrew J, D. Tek bir oryantasyonun konturları ile erken görsel deneyimin kedilerde fizyolojik ve davranışsal bir çalışması, J. Physiol. Londra., 1977, 265, 615-636.
Brown J., Lewis V., Brown M., Horn G., Bowes J. B. İki ilacın (diazepam ve lorazepam) neden olduğu geçici amneziler ile organik kökenli amnezi arasında bir karşılaştırma, Neurophysiologia, 1982, 20, 55-70.
Brown J., Brown MW, Bowes JB Lorazepam'ın unutma oranı, semantik bellekten geri çağırma ve el becerisi üzerindeki etkisi, Neuropsychologia,. 1983, 21, 501-512.
Kahverengi MW Davranışsal olarak eğitilmiş maymunların hipokampal bölgesinden kaydedilen tek birimlerin tepkisi üzerinde bağlamın etkisi. In: Neuronal plasti city and memory function (eds. C. Ajmone Marson ve H. Matthies), 1982,. pp. 557-573. Raven Press, New York.
Brown MW, Horn G. Eğitilmiş maymunların kalça pokampal bölgesinden kaydedilen bağlama bağımlı nöronal tepkiler , J. Physiol. Londra., 1978, 282, 15-16P.
Bruce C., Desimone R., Gross CG Makağın üst temporal sulkusundaki çok duyusal bir alandaki nöronların görsel özellikleri, J. Neurophysiol., 1981, 46, 369-384.
Cherfas JJ, Scott A. Evlat damgalamanın süreksiz tersine çevrilmesi, Anim. Behav., 1981, 29, 301.
Cohen NJ, Squire LR Amnezide örüntü analizi becerisinin korunmuş öğrenmesi ve akılda tutulması: nasıl olduğunu bilmenin ve neyi bilmenin ayrışması, Science, 1980, 210, 207-209.
De Renzi E. Fokal neokortikal hasarın ardından hafıza bozuklukları, Phil. Trans. R. Soc., 1982, B298, 73-83.
Dickinson A. Çağdaş hayvan öğrenme teorisi. Cambridge Üniversitesi Yayınları. Cambridge, 1980.
Divae Kosmal A., Björklund A., Lindvall O. Yaban turpu peroksidaz tekniği ile ortaya çıkarılan sıçandaki prefrontal kortekse subkortikal projeksiyonlar, Neuroscience, 1978, 3, 785-796.
Freeman DN, Morg E. Görme keskinliği gelişimi, kedi yavrularında hassas dönemle çakışır, Nature, Londra., 1975, 254, 614-615.
Gaffan D., Saunders RC, Gaffan EA, Harrison S., Shields C., Owen MJ Maymunlarda çağrışımsal hafıza üzerindeki forniks transeksiyonlarının etkileri: öğrenilmiş eylemde hipokampusun rolü, Quart. J. exp. PsychoL, 1984, 36B, 173-221.
Gören CC, Sarty M., Wu PYK Görsel takip ve yüz benzeri uyaranların yeni doğan bebekler tarafından örüntü ayrımı, Pediatrics, Springfield, 1975, 56, 544-549.
Guiton P. Damgalama kavramıyla ilgili olarak evcil kümes hayvanlarında cinsel tepkilerin gelişimi, Symp. hayvanat bahçesi sos. Londra., 1962, 8, 227-234.
Guiton R. Brown Leghorn'da erken deneyim ve cinsel nesne seçimi, Anim.Behav., 1966, 14, 534-538.
Hagbarth KE, Kugelberg E. İnsan karın derisi refleksinin plastisitesi^ Brain, 1958, 81, 305-318.
Nöcaen ti. Frontal lob tümörleri ile ilişkili zihinsel semptomlar. In: Frontal granüler korteks ve davranış (ed. JM. Warren ve A. Akert), 1964, s. 335-352, McGraw-Hill, New York.
Hinde RA Damgalama sorununun bazı yönleri, Symp. hayvanat bahçesi sos. Londra., 1962, 8, 129-138.
Hinde RA Etolojisi: doğası ve diğer Bilimlerle ilişkileri. 1982. Fontana, Londra.
Hinde RA, Stevenson-Hinde J. (ed.). Öğrenme üzerindeki kısıtlamalar, 1973, Academic Press, Londra.
Hirsch HVB, Spinelli DN Görsel deneyim, kedilerde yatay ve dikey olarak yönlendirilmiş alıcı alanların dağılımını değiştirir, Science, 1970, 168, 869-871.
Horn G. Cat striatum korteksindeki tek birimlerin düzenli dürtü aktivitesi, Nature, Londra., 1962, 194, 1084-1085.
Horn G. Nöronal aktivitedeki değişiklikler ve bunların davranışla ilişkisi. İçinde: Nöral aktivite ve davranışta kısa vadeli değişiklikler. (editörler G. Horn ve R.A. Hinde), 1970, s. 567-606. Cambridge University Press, Cambridge.
Horn G., Rose SPR, Bateson PPG Merkezi sinir sisteminde deneyim ve plastisite, Science, 1973, 181, 506-514.
Hubel DH, Wiesel TN Alıcı alanlar, binoküler etkileşim ve kedinin görsel korteksindeki işlevsel mimari, J. Physiol. Londra., 1962, 160, 106-154.
Hubel DH, Wiesel TN Yavru kedilerde tek taraflı göz kapatmanın fizyolojik etkilerine duyarlılık dönemi, J. Physiol. Londra., 1970, 206, 419-436.
Huppert FA, Plercy M. Unutkanlar ve kontrol denekleri tarafından yakınlık ve sıklık kararlarında iz gücünün rolü, Quart. J. exp. Psychol., 1978, 30, 346-354.
İversen SD Hipokampal lezyonlar hayvanlarda amnezi yapar mı? Int. Rev. Neurobiol., 1976, 19, 1-49.
versen SD Hayvanlarda beyin lezyonları ve hafıza: yeniden değerlendirme. İçinde: Belleğin fizyolojik temeli (ed. JA Deutsch), 1983, s. 139-198, Academic Press, New York.
Jacklet JW Sinir sistemindeki endojen osilatörler tarafından sirkadiyen zamanlama: hücresel mekanizmalara doğru, Biol. Bull., 1981, 160, 199-227.
James WJ Psikolojinin ilkeleri. Henry Holt, New York, [Yeniden basılmıştır (1950) Dover Publications, New York], 1890.
Kamil AC Nektarla beslenen bir kuş olan Amakihi (Loxops virens), J. comp. tarafından sistematik yiyecek arama. fizik. Psychol., 1978, 92, 388-396.
Kratz KE, Lehmkuhle S. Tek gözlü yoksun kedilerde yoksun olmayan gözün çıkarılmasından sonra mekansal kontrast duyarlılığı, Behav. Brain Res., 1983, 7, 261-266.
Lorenz K. Der Kumpan in der Umwelt des Vogels, J. Orn., Lpz., 1935, 83, 137-213, 289-413.
Lorenz K. Kuşların dünyasında yol arkadaşı, Auk, 1937, 54, 245-273.
Mabille E., Pitres A. Sur un cas d'amnesie de fixation postapoplectique ayanl persit6 vingt-trois ans, Revue Med., 1913, 33, 257-279.
Mackintosh NJ Koşullandırma çeşitleri. In: Öğrenme ve belleğin nörobiyolojisi üzerine ikinci konferans (editörler G. Lynch, JL McGaugh ve NM Weinberger), 1986, s. 335-350, Guilford Press, New York.
Maudsley HI Aklın Fizyolojisi, 1876, Londra.
Marler P. Song öğrenme: öğrenme sürecinde doğuştan gelen tür farklılıkları. İçinde: Öğrenmenin biyolojisi (eds. P. Marler ve HS Terrace), 1984, s. 289-309, Dahlem Konferenzen, Springer, W. Berlin.
Hauta WJH, Karten HJ Omurgalı beyninin genel bir profili, serebral korteksin ataları üzerinde yan ışıklar. İçinde: Nörobilimler, ikinci çalışma programı (ed. F.O. Schmitt), 1970, s. 7-26. Rockefeller University Press, New York.
Pay ne JK t Horn G., Brown M. no. Civciv beyninin görsel bir projeksiyon alanındaki yanıt verebilirliğin değiştirilebilirliği: görsel deneyim, ilgili birkaç faktörden yalnızca biridir, Davranış. Brain Res., 1984, 13, 163-172.
Perrett D. 1., Rolls ET t Caan no. Maymun temporal korteksindeki yüzlere yanıt veren görsel nöronlar, Exp. Brain Res., 1982" 47, 229-238.
Pettigrew JD t Konishi M. Bamkuşunun (Tyto alba) görsel Wulst'unda yönelim ve dürbün eşitsizliği için seçici nöronlar, Science, 1976a, 193, 675-678.
Pettigrew JD, Konishi M. Baykuşun görselindeki binoküler nöronlar üzerinde monoküler yoksunluğun etkisi Wulst, Nature, Londra., 1976b, 264, 753-754.
Rolls ET, Perret D. Caan A. no., Wilson FA no. Görsel tanıma ile ilgili nöronal tepkiler, Brain, 1982, 105, 611-646.
Ryle G. Zihin kavramı, 1949. Hutchinson, Londra.
Salzen EA Evcil civcivlerin davranışında deneyim, uyaran özellikleri ve eksojen androjen etkileşimi, Behavior, 1966, 26, 286-322 .
Salzen EA, Meyer C. Baskının tersine çevrilebilirliği, J. comp. filol. Psychol., 1968, 66, 269-275.
Schutz F. Sexuelle Pragung bei Anatiden, Z. Tierpsychol., 1965, 22, 50-103.
Seligman MEP, Hager JL Biyolojik öğrenme sınırları, 1972. Appleton—Century—Crofts, New York.
Sluckin No., Salzen EA damgalama ve algısal öğrenme, Quart, J. exp. Psychol., 1961, 13, 65-77.
Stechler G. Doğum sırasında ilacın yenidoğan dikkati üzerindeki etkisi, Science, 1965, 144, 315-317.
Strumwasser F. Davranışın geçici organizasyonundaki sinirsel ve hümoral faktörler, The Physiologist, 1973, 16, 9-42.
Tulving E. Olaysal ve anlamsal bellek. İçinde: Hafıza organizasyonu (editörler E. Tulving ve W. Donaldson), 1972, s. 382-403, Academic Press, New York.
Vinogradova O. S. Bilgi Kaydı ve limbik sistem. İçinde: Nöral aktivite ve davranışta kısa süreli değişiklikler (eds. G. Horn ve R.A. Hinde), 1970, s. 95-140. Cambridge University Press, Cambridge.
Walsh K. hayır. Nöropsikoloji: klinik bir yaklaşım, 1978, Churchill Livingstone, Edinburgh.
Warrington EK, Weiskrantz L. Amnezi: Bir kopukluk sendromu mu? Nöropsikoloji, 1982, 20, 233-248.
Weiskrantz L. Amnezinin deneysel çalışmaları. In: Amnesia (editörler CWM Whitty ve OL Zangwill), 1966, s. 1-35, Butterworth, Londra.
Weiskrantz L. Hafıza kaybı çalışmalarının karşılaştırmalı yönleri, Phil. Trans. R. Soc., 1982, B298, 97-109.
Weiskrantz L. İnsan amnezik sendromunun sorunları ve teorileri üzerine. İçinde: Öğrenme ve hafızanın nörobiyolojisi üzerine ikinci konferans (editörler G. Lynch, JL McGaugh ve NM Weinberger), 1986, s. 380-415, Guilford Press, New York.
Whitty C. hayır. M., Lishman hayır. A. Serebral hastalıkta amnezi. In: Amnesia (editörler, CWM Whitty ve OL Zangwill), 1966, s. 36-76. Butterworth, Londra.
Wiesel TN, Hubel DH Tek gözde Görmeden yoksun bırakılan yavru kedilerin çizgili korteksindeki tek hücreli yanıtlar, J. Neurophysiol., 1963, 26, 1003-1017.
Wilson P. Evcil civcivlerde görsel hiperstriatumun organizasyonu. II. Tek birimlerin alıcı alan özellikleri, Brain Res., 1980, 188, 333-345.
Zola-Morgan S., Squire LR Medial temporal lezyonları olan maymunlarda korunmuş öğrenme : motor ve bilişsel becerilerin korunması, J. Neurosci., 1984, 4, 1072-1085.
EK
KUŞLARIN BEYNİ YAPISININ, ÖZELLİKLE EVCİL TAVUKLARININ YAPISI
Bu makalenin amacı, bu kitapta anlatılan deneysel araştırmaların yapısal temellerini göstermektir. Bu nedenle, açıklamamızın kendine özgü bir özelliği vardır: görsel yollara, beyin sapı projeksiyonlarına ve memelilerin ve kuşların beyinlerinin karşılaştırmalarına el kitaplarında alışılageldiğinden daha fazla yer ayrılmıştır. Buna ek olarak, bu konuyla ilgili çok sayıda mükemmel çalışma olduğundan, bu konunun ayrıntılı sunumu gereksizdir (Kappers ve diğerleri, 1936; Pearson, 1972; Cohen ve Karten, 1974; Macphail, 1982).
Belirli bir tür davranışın sinirsel mekanizmalarını anlamak için, onu sağlayan yapıları anlamak gerekir. Araştırmamızı evcil tavukların beyni üzerinde gerçekleştirdik. Nasıl görünüyor, ana departmanları ve aralarındaki bağlantılar neler?
S.1. Tavuk beyninin genel tanımı
İki günlük bir civcivin beyni 1 ml'den biraz fazla su taşır; uzunluğu yaklaşık 20 mm ve en büyük genişliği 14 mm'dir. İlk bakışta bu beyin diğer pek çok omurgalının beyinleriyle aynı görünmektedir (Şekil A.1). Serebral hemisferler , beyincik ve beyin sapı görülebilir. Omurgalılarda olduğu gibi, civcivin beyninin makroskobik organizasyonu, embriyolojik gelişimi bilgisi ile anlaşılması kolaydır.
Merkezi sinir sistemi, içi boş bir silindir olan nöral tüpten gelişir (Şekil P.2,L). Tüp kısmının önündeki merkezi boşluk daha sonra yetişkin beyninin ventriküler sistemini oluşturur ve onun maddesi tüpün duvarlarından oluşur. Nöronlar ve destekleyici glial hücreler, tüpü kaplayan epitelden türetilir. Nöronlar dış yüzeye göç eder , bu da gelecekteki beynin duvarlarının kalınlaşmasına, yüzeyde çıkıntılar olarak görülebilen kümeler oluşturmasına veya tüpün yüzeyini kaplayan katmanlar oluşturmasına yol açar.
Gelişimin çok erken bir aşamasında, tüpün ön ucunda göze çarpan üç kalınlaşma vardır (Şekil A.2, B). Daha sonra beynin ana bölümleri onlardan oluşur. Arka balondan
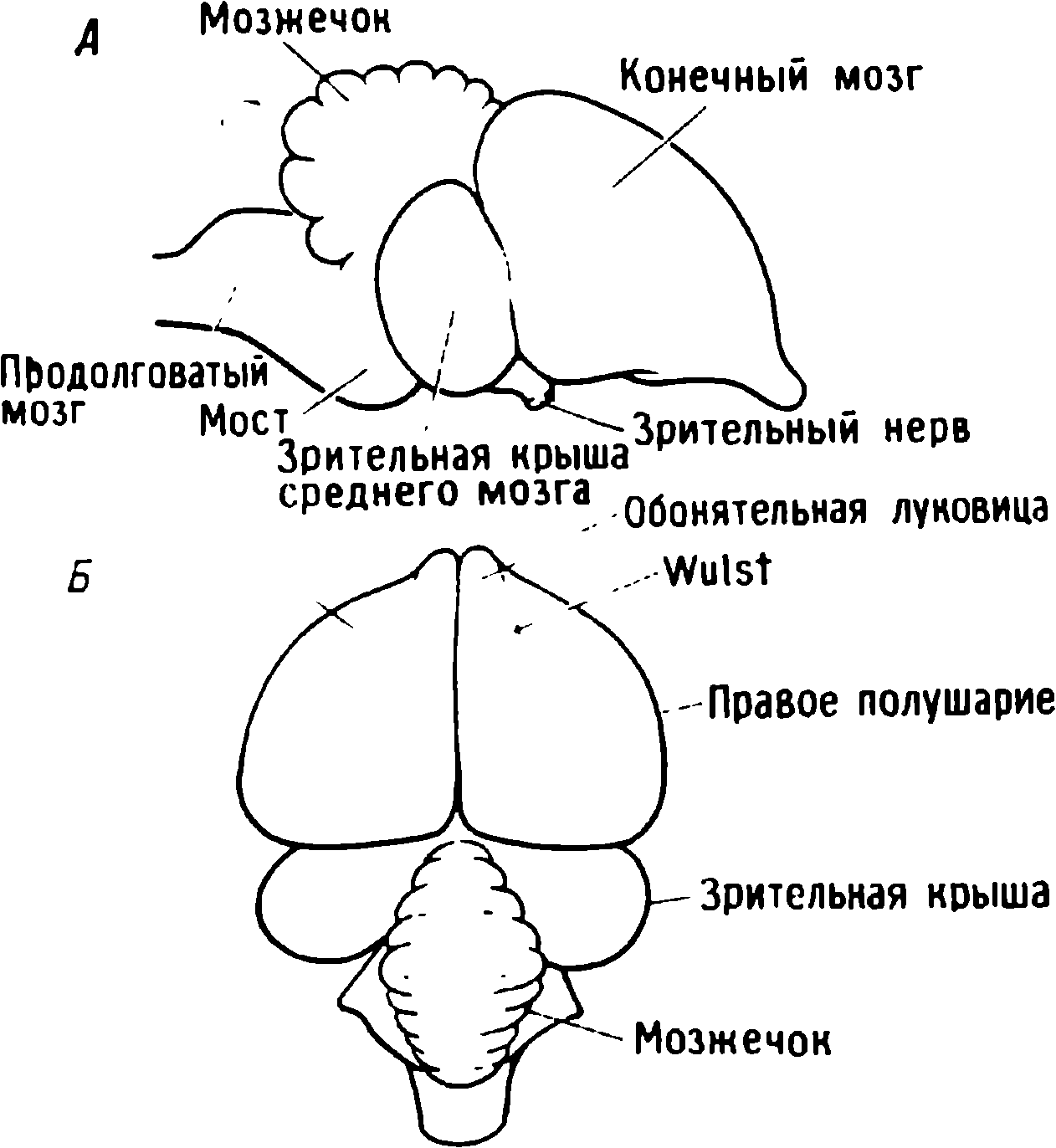

5 mm
Resim: S.1. İki günlük bir tavuğun beyni; yandan görünüm (4), üst (B) ve alt (C).
arka beyin önden - ön beyinden ve diensefalondan - orta beyinden oluşur. Ön beyin iki ana bölümden oluşacaktır - diensefalon (diensefalon ) ve telensefalon (telensefalon). Telensefalonun en karakteristik özelliği hemisferlerin varlığıdır. Nöral tüpün ön mesanesinin çıkıntıları olarak ortaya çıkarlar (Şekil A.2, C). Bu çıkıntıların duvarları serebral hemisferlerin maddesini oluşturur ve boşluklar lateral ventriküllere dönüşür. Gelişmekte olan beyin, orijinal nöral tüpün basit şeklini değiştiren bükülür. Bu fenomenler , kuluçkanın 8. gününde embriyoda bile meydana gelir (Şekil P.2, D).
Serebellum ilk önce arka mesanenin dorsal kısmında çoğalan hücrelerin iki taraflı bir koleksiyonu olarak görünür.
140
dördüncü ventrikül
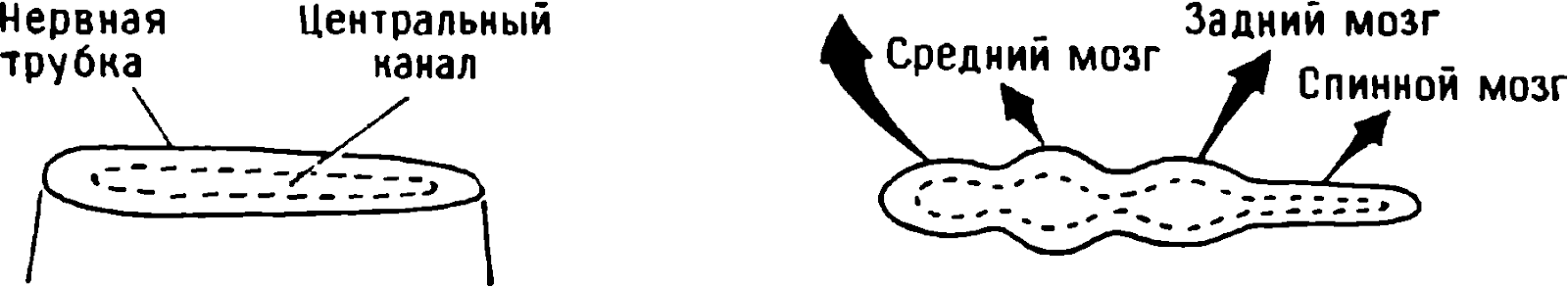
Omurilik
İki hücre kitlesi orta hatta birleşerek tek bir ikili yapı olan serebellumu oluşturur. Benzer şekilde hücreler, orta beynin oluştuğu ara vezikülde çoğalır. Görsel sistemle olan bağlantılarına göre, orta beynin bir bölümü görsel çatı olarak adlandırılır (Şekil A.1).
Nöral tüpün boşluğunun gelişmekte olan telensefalonun arkasındaki kısmından üçüncü ventrikül oluşur. Bu ventrikül , duvarlarını oluşturan madde ile birlikte diensefalonu oluşturur (Şekil A.2, C ). Hücrelerin bu yapılardaki birikimleri ayrı çekirdekler oluşturur. Bu çekirdeklerin iki ana birliği , sırasıyla talamus ve hipotalamusu oluşturur (Şekil P.2, D).
S.2. Telensefalonun iç yapısı
Makroskopik düzeyde, kuşların beyinleri, yarımküreleri kıvrımlardan yoksun olan memelilerin beyinlerine genel bir benzerlik gösterir. Bununla birlikte, bu iki hayvan sınıfının ön beyin yarım kürelerinin iç yapısı çarpıcı biçimde farklıdır. Ön kısımda, Memelilerde hemisferlerin dış tabakası serebral korteks tarafından oluşturulur. Boyanmamış bölümlerde, katmanlı nöronlardan oluşan korteks gri görünür. Hücre katmanlarının altında, çoğu miyelinli olan birçok akson vardır; Bu alanda nöronların gövdeleri yoktur. Miyelinli aksonların kütlesi , subkortikal maddeye parlak beyaz bir renk verir. Kuşların beyninde gri ve beyaz madde çok net bir şekilde ayrılmamıştır. Bir tavuğun beyin bölümleri, hücre gövdelerini boyayan boyalarla boyandığında, bu gövdelerin ön beyin boyunca dağıldığı görülür ve bu, memelilerin serebral korteksine hiç benzemez .
Civciv beyni, yanal ve rostro-kaudal yönlerde boyutları farklı olan temel olarak hücre katmanlarından oluşur (Şekil P.3). Bazı yerlerde bu katmanlar , ektostriatum gibi hücre kümelerini sarar (Şekil P.3, B). Her iki serebral hemisferin ön yarısındaki en yüzeysel üç katman , orta hatta yakın bir tepe oluşturur. Bu nispeten geniş yastık şeklindeki yükselti, Wulst olarak bilinir (Şekil A.1, B). Wulst, ek ve dorsal bir hiperstriatum (hyperstriatum accessorium ve hyperstriatum dorsale) ile bunların arasına sıkıştırılmış bir ara çekirdek veya bir ara hiperstriatum (hyperstriatum intercalatus) içerir. Beynin ön kısmında dorsal hiperstriatumun altında, ancak arka kısmında lateral ventrikülün hemen altında ventral hiperstriatum bulunur (hyperstriatum ventrale ; Şekil P.Z, E). Bu yapı, ön beyin - neostriatum'un (neostriatum) en büyük hücre tabakasının üzerinde yer alır. Ectostriatum ve archistriatum, neostriatal hücreler tarafından değişen derecelerde çevrelenmiş kabaca yuvarlak hücre kümeleridir. Ön beyin tabanında yer alan yükselen paleostriatum (raieostriatum boostatum), ilkel paleostriatum (raieostriatum primitivum) üzerinde bir başlık oluşturur (Şekil P.3, C, D). Ventrikülün medialinde hipokampus ve septal çekirdekler bulunur.
Kuşların ve memelilerin beyinleri arasındaki önemli yapısal farklılıklara daha önce işaret etmiştik. Kuş beyninin şu ya da bu bölümünün, memeli beyninin bazı bölümleriyle homolog olduğunu varsaymak doğru mudur? Yaklaşımlardan birine göre, memelilerin ve kuşların yetişkin beyinlerinin bazı bölgeleri aynı embriyonik kökene sahipse, bu bölgelerin homolog olduğu varsayılabilir (örneğin bkz. Kuhlenbeck, 1938; Kâllen, 1962; Benowitz, 1980). Serebral veziküllerin oluşumundan kısa bir süre sonra, yan duvarlarının iç yüzeyinde bir oluk oluşur. 21*

Resim: PZ İki günlük bir tavuğun ön beyin bölümlerinin şeması. Üstteki dört görüntü beynin ön bölümleridir. Şemalar A ve B , ön beynin rostral üçte biri seviyesindeki bölümleri temsil eder ( A seviyesi , B'nin önündedir ). Şema C , orta üçlü seviyesinde bir bölümü ve önbeynin arka üçte biri seviyesinde Şema D'yi temsil eder. E. Beynin uzunlamasına kesiti orta hatta biraz yanal. 1 - ek hiperstriatum; 2 - ara hiperstriatum; 3 - dorsal hiperstriatum; 4 - ventral hiperstriatum; 5 - neostriatum; 6 - bazal çekirdek; 7 - para koku alanı; 8 - ektotriyum; 9 - yükseltilmiş paleostriatum ; 10 - hipokamp; 11 - parahipokampal bölge; 12 - ventrikül; 13 - dorso-lateral kortikoid bölgesi; 14 - septal çekirdekler; 15 - arşiv ; 16 - ilkel paleostriatum; 17 - görsel çatı.
önden arkaya uzanan, her baloncuğu dorsal ve ventral bileşenlere bölen (Şekil A.4, L). Serebral veziküllerin farklı bölümlerinin nihai gelişiminin sonucu, Şekil 1'de tavuk için gösterilmiştir. P.4, B. Memelilerde, kabarcıkların yan duvarının ventral bileşeni, striatum veya "striatum" (corpus striatum) olarak bilinen bir yapı kompleksi oluşturur. Sırt bölümü, serebral korteksin oluştuğu palyumu oluşturur. Dorsal bölgenin medial kısmı hipokampal korteksi oluşturur.


Resim: S.4. Tavuğun ön beyninin yarım kürelerinin gelişim aşamalarının şeması. A. Yaklaşık olarak inkübasyonun 4. gününde embriyonun iki serebral vezikülü. Kesikli çizgi, kabarcıkların dorsal tarafı (D. s.) ve ventral tarafı (V. s.) arasındaki sınırı işaretler. B. Sağ yarım küre. İnkübasyonun 7. gününde lateral duvar hücreleri çoğalmaya başlar ve ventriküle doğru bir çıkıntı oluşturur. Fi ve D , embriyonik anlajın geliştiği kesin yapıları gösterir . Bu diyagramlar, iki günlük bir civcivin beyninin ön (C) ve arka (D) bölümlerinin kesitlerini göstermektedir. 1 - palyum; 2 - hipokamp; 3 - parahipokampal bölge; 4 - Vulst; 5 - ventral hiperstriatum; 6 - neostriatum; 7 - ektotriyum; 8 - anterior archistriatum (archistriatum'un ara ve dorsal bileşenleri , beyin mesanesinin dorsal kısmından da oluşturulabilir); 9 - medial archistriatum (archistriatum'un arka çekirdeği , beyin mesanesinin ventral kısmından da oluşturulabilir); 10 - bazal çekirdek; // - yükselen paleostriatum; 12 - ilkel paleostriatum; 13 - septal çekirdekler; 14 - ek hiperstriatum; 15 - ara hiperstriatum; 16 - dorsal hiperstriatum; 17 - ventrikül. (Kuhlenbeck, 1938; Kaien, 1962; Karten, 1969; Nauta ve Karten, 1970; Benowitz, 1980.)
Beynin ontogenezi tarafından yönlendirilirsek, kuşlarda ve memelilerde bazı bölümler homolog olarak kabul edilebilir . Özellikle, memeli neokorteksinin , hiperstriatal kompleksin (dorsal ve ventral hiperstriatum dahil) birkaç bileşenine ve neostriatum ve ectostriatum kombinasyonuna homolog olduğu görülmektedir. Kuşlarda ve memelilerde hipokampus ve parahipokampal bölge de homolog olabilir. Olası karşılaştırmaların bu listesi ayrıntılı değildir (Kâllen, 1962; Benowitz, 1980), ancak bu tür karşılaştırmaların işlevsel önemi belirsizliğini koruyor. Bir dizi embriyolojik kritere göre iki yapı homolog görünüyorsa, bu mutlaka işlevlerinin benzerliği anlamına gelmez. Ancak bir hayvan sınıfında verilen bir yapının işlevi biliniyorsa, başka bir sınıftaki benzerinin de benzer bir işleve sahip olup olmadığının bulunması şüphesiz gereklidir.
PZ Sensör sistemleri
değişikliklerin sinir sisteminde neden olduğu bazı sinyaller ön beyne ulaşır. Bu sinyallerin iletim yolları yoğun araştırmalara konu olmuştur. Modern bilgi, büyük ölçüde , geçmişin sonunda ve yüzyılımızın başında nöroanatomistlerin zahmetli ve çoğu zaman parlak araştırmalarına dayanmaktadır. Bu çalışmalar, duyusal sistemleri incelemek için yeni anatomik ve fizyolojik yöntemler sayesinde daha da geliştirilmiştir . Genellikle bu çalışmalar güvercinler (Columba Іѵіа) üzerinde yapılmaktadır. Bu türdeki beynin temel organizasyonu, incelenen diğer kuş türlerinin beyin yapısına benzer (Pearson, 1972). Ancak , farklı kuş takımlarında ve hatta aynı takım içindeki farklı cinslerde beynin sito-mimari yapısında önemli farklılıklar olduğu unutulmamalıdır (bkz. örneğin Stingelin, 1958; Cobb, 1960a, b). Bu farklılıklar , beynin çeşitli bölümlerinin ve yollarının ultrastrüktürel ve fizyolojik özelliklerinde kendini gösterebilir . Aşağıda bu sınırlamayı her zaman dikkate almazsak, bu, kuşların karşılaştırmalı fizyolojisine değil, öncelikle öğrenmenin nöral mekanizmalarına ilgi duyan okuyucular tarafından metnin okunmasını kolaylaştırmak için yapılır. beyin.
Madde 3.1. görsel sistem
Güvercinlerde, optik sinir lifleri tamamen optik kiazma bölgesinde kesişir (Cowan ve diğerleri, 1961). Sonuç olarak, sol retinadan gelen sinyaller beyin sapının sağ tarafındaki yapılara gider. Optik sinirdeki aksonların iki ana hedefi optik çatı ve talamustur (Cowan ve diğerleri, 1961). Görsel çatıdan geçen yola tektofugal, talamustan gelen yola talamofugal denir.
Tektofugal yolun lifleri görsel çatıdan çıkar ve yuvarlak çekirdeğe (n. rotundus) yansıtılır (Şekil A.5, L). Geçişten sonra bu yol ektostriatumun orta kısmında son bulur (Karten ve Hodos, 1970). ektostriatum içinde
Resim: S.5. Görsel sistemin ana yükselen bağlantılarının şeması. A. Retinanın sadece kontralateral hemisfere çıkıntı yaptığı tektofugal yol . B. Talamofugal yol. 1 - yükselen paleostriatum; 2 - periectostriatal kemer; 3 - ventral hiperstriatum; 4 - dorsal hiperstriatum; 5 - ara hiperstriatum; 6 - ek hiperstriatum; 7 - ektotriyum; 8 - neostriatum; 9 - yuvarlak çekirdek; / 0 - görsel çatı; 11 - optik kiazma; 12 - sağ optik sinir; 13 - sol optik sinir; 14 - dorsal supraoptik komisyon; 15 - talamusun ana görsel çekirdeği. Teraziye uyulmuyor. (Karten. 1969, değişikliklerle.)
sırayla kendisini kısmen çevreleyen hücrelere, sözde periektostriatal kuşağa bağlantılar gönderir.
Talamofugal sistem ayrıca bilgiyi telensefalona iletir (Şekil A.5, B). Optik sinir lifleri , talamusun çok çekirdekli kompleksinde son bulur. Bu komplekse talamusun ana görsel çekirdeği denir. Hücrelerinin aksonları, önbeynin ipsi- ve kontralateral hemisferlerine yansıtılır. İpsilateral projeksiyon, görsel wulst'un üç bileşeninde de sona erer . Kontralateral izdüşüm dorsal supraoptik komissür yoluyla yükselir ( Hunt ve Webster, 1972; Karten ve diğerleri, 1973). Peçeli baykuşta (Tyto alba), görsel Wulst hücrelerinin çoğu her iki gözün uyarılmasına tepki verirken (Pettigrew ve Konishi, 1976), tavukta ise hücrelerin sadece küçük bir kısmı bu tür özelliklere sahiptir (Brown ve Hoth, 1979). ; Wilson, 1980). Aksesuar hiperstriatumda binoküler yakınsama sağlayan anatomik yol bilinmemektedir. Bununla birlikte, Wulst'un derin katmanları ikili yollar alır. Bu katmanlar, aksesuar hiperstriatum üzerine yansıtılır (Bradley ve diğerleri, 1985) ve ona gerekli sinyalleri iletebilir. Böylece talamofugal sistem , sağ ve sol gözlerden her yarımkürenin Wulst'una oldukça organize bir bilgi akışı sağlar . Buna karşılık, tektofugal sistem, bilgiyi hemisfere yalnızca kontralateral gözden iletir.
Yakın zamanda evcil tavuklarda küçük, çaprazlanmamış bir görsel yol tanımlanmıştır (O'Leary ve diğerleri, 1983). Retinadan gelen lifler ipsilateral talamusa ve orta beyne gider. Bu sistemin işlevleri henüz araştırılmamıştır.
Wulst'a ek olarak , ön beynin birkaç başka bölgesi de görsel bilgi alır. Güvercinlerde (Parker ve Delius, 1972) ve gri kaz Anser anser'de (Felix ve diğerleri, 1983) neostriatumun arka kısmında görsel uyarılmış potansiyeller kaydedilmiştir . Periectostriatal kuşağın nöronlarının görsel sinyalleri bu alana iletmesi olasıdır (Şekil P.5, L) . Bu nöronlar aynı zamanda archistriatum'un dorsal kısmındaki hücre gruplarına da yansır (Ritchie ve Cohen, 1977).
Son zamanlarda açıklanan yol , optik çatının ventral hiperstriatum ile doğrudan bağlantısı olabilir (Bradley ve Hoot, 1978; Bradley ve diğerleri, 1985). Görsel bilginin ön beyne ulaşabileceği bu yol , Bölüm 1'de ayrıntılı olarak tartışılmaktadır. 11.1.1.
Madde 3.2. işitme sistemi
İşitsel bilginin kulaktan ön beynin karşı yarımküresine iletilmesinin ana yolu, Şekil 1'de gösterilmektedir. S.6. İç kulağın kokleasından gelen sinyaller, medulla oblongata'nın iki çekirdeğine iletilir (Boord ve Rasmussen, 1963). Bu çekirdeklerin hücrelerinden gönderilen aksonlar , beyin sapının karşı tarafında yükselen yanal döngü adı verilen bir yolun parçasını oluşturur . Bu liflerin çoğu orta beynin dorsolateral çekirdeğinde son bulur (Şekil P.6, 5). Daha sonra bu yol talamusun oval çekirdeğine kadar devam eder ve oradan geçerek L alanı olarak adlandırılan arka neostriatum bölgesine gider (Karten, 1967, 1968; Bonke ve diğerleri, 1979). Bu, iç kulaktan ön beyne giden doğrudan yoldur. Gerçekte , işitsel yollar, Şekil 2'de tasvir edilenlerden çok daha karmaşıktır. S.6. Bu şekilde gösterilen iki medulla oblongata çekirdeği sadece ipsilateral kulaktan girdi alırken (Erulkar, 1955), diğer beyin sapı çekirdekleri her iki kulaktan da girdi alır (Erulkar, 1955; Board, 1968). Peçeli baykuşta, sağ ve sol işitsel alanlardan gelen bilgiler, orta beynin dorsolateral çekirdeğinin bir kısmının tek hücrelerinde birleşir . Bu bilgi, işitsel alanın doğru bir haritasını oluşturmak için kullanılır (Konishi ve Knudsen, 1983). işitsel pu-

Resim: S.6. Yükselen işitsel yolların şeması. İşitme sinirinin (9) koklear kısmındaki primer afferent lifler , medulla oblongata'da (10) iki işitsel çekirdekte (8) son bulur. Postsinaptik nöronları , biri orta beyinde olan diğer çekirdeklere projekte olur; ona giden yollar yanal döngüye (1) gider. Orta beynin (3) dorsolateral çekirdeği olan bu çekirdek, görsel çatının (2) altında yer alır ve oval çekirdeğe (4) çıkıntı yapar . Bu çekirdekte geçiş yaptıktan sonra, yol L alanına ve ardından ventral hiperstriatumun kaudal kısmına gider (7). Yalnızca kontralateral yol gösterilir. 5 - arşiv; ey mide Teraziye uyulmuyor. (Karten, 1968; Board, 1969; Bonke ve diğerleri, 1979.)
belirli hücrelerin ayrı ses frekanslarına tepki vermesi için tonotopik olarak düzenlenirler (Stopp ve Whitefield, 1961; Zaretsky ve Konishi, 1976). İki işlev, ses lokalizasyonu ve frekans analizi, anatomik olarak ayrılmış görünmektedir (Sachs ve Sinnott , 1978; Knudsen ve Konishi, 1978).
L alanı ayrıca bir tonotopik organizasyona sahiptir (Zaretsky ve Konishi, 1976; Scheich ve diğerleri, 1979a; Scheich, 1983). Beç tavuğu (Nutnida meleagris) ve sığırcıkta (Sturnus vulgaris), L alanındaki tek hücreler karmaşık ses kombinasyonlarına yanıt verir. Bazı nöronlar, türe özgü seslerin belirli bileşenleri için oldukça seçicidir (Leppelsack ve Vogt, 1976; Scheich ve diğerleri, 1979b). L alanı , beç tavuğunda gösterildiği gibi ventral hiperstriatumun kaudal kısmına çıkıntı yapar ( Bonke ve diğerleri, 1979).
Son yıllarda, L alanında bitmeyen telensefalona giden işitsel yolların varlığına ilişkin veriler birikmiştir. Tavuklar da dahil olmak üzere birçok kuş türünde neostriatumun ön kısmında işitsel uyarılmış potansiyeller kaydedilmiştir (Harman ve Phillips, 1967) ve güvercinler (Ilyichev ve diğerleri, 1970; Delius ve diğerleri, 1979). Görünüşe göre, bu yol neostriatumun nükleus bazalis yakınındaki alanına götürür (Şekil P.3, E). Bu alanda kaydedilen uyarılmış potansiyellerin , muhtemelen gagada bulunan dokunma veya titreşim alıcılarının kasıtsız uyarılmasından kaynaklanan eserler olduğu varsayılmıştır (Karten, 1968). Delius ve diğerleri ( 1979) güvercindeki çift taraflı koklear yıkımın bu uyarılmış potansiyelleri ortadan kaldırdığını gösterdiğinden, bu olası değildir . Ek olarak, Kirsch ve diğerleri ( 1980), uyanık sığırcıklarda neostriatumun bu kısmındaki bireysel hücrelerin aktivitesini kaydetti ve farklı hücrelerin farklı ses uyaranlarına tepki verdiğini buldu . Bu iki deney grubu, işitsel bölgenin aslında neostriatumun ön kısmında yer aldığını göstermektedir. Bu sistemin anatomik yolları bilinmemekle birlikte ön beyne çok hızlı iletim sağlarlar. Bu hız o kadar yüksektir ki, büyük olasılıkla, bu yol talamusta geçiş yapmadan ön beyne ulaşır.
Bir bütün olarak işitsel yolların organizasyonu birçok kuş türünde ortaktır. Bununla birlikte, ince organizasyonunun değişmez olması ve örneğin ötücü kuşların işitsel sisteminin diğer kuş türlerininkiyle tamamen aynı olması şaşırtıcı olurdu.
Madde 3.3. Somatosensoriyel sistem
Yüz yüzeyinden gelen sinyallerin hareket ettiği yollar biraz ayrıntılı olarak açıklanmaktadır, ancak omurilikte somatosensoriyel sinyallerin iletimi hakkında çok az şey bilinmektedir. Çeşitli çalışmalarda, güvercinlerin telensefalonlarında somatosensoriyel uyarılmış potansiyeller kaydedilmiştir (Erulgar, 1955; Delius ve Bennetto, 1972). Tepkiler , cilt sinirlerinin doğrudan uyarılmasının yanı sıra vücut yüzeyinin dokunsal uyarılmasıyla ortaya çıktı (Delius ve Bennetto, 1972). Her iki durumda da, baş da dahil olmak üzere vücudun kontralateral tarafının uyarılmasına yanıtlar, orta hatta yakın neostriatumun kaudal kısmında ortaya çıktı. Gri kazdaki somatosensoriyel yolların çalışmasında da benzer sonuçlar elde edilmiştir (Felix ve diğerleri, 1983). Güvercinlerde neostriatumun kaudal kısmının somatosensoriyel projeksiyon alanı, L. Delius ve Bennetto (Delius ve Bennetto, 1972 ) alanının önünde ve üzerinde yer alır . yaklaşık olarak Şekil 2'de gösterilen ara hiperstriatuma karşılık gelen bölge . GEZ, A.
Yüz yüzeyinden duyusal bilgileri taşıyan yollardan biri, yıllar önce Wallenberg (Wallenberg, 1903) tarafından tanımlanmıştır. Bu bölgeyi innerve eden birincil duyu lifleri beyin sapında son bulur. Postsinaptik nöronlar , talamusu atlayarak doğrudan ön beyine projekte olur. Bu sistemin birincil afferent lifleri, beşinci kranial sinirin bir parçasıdır. Buna göre Wallenberg, ön beynin bu izdüşümünü quintofrontal yol olarak adlandırdı. Her iki taraftan lifler içeren bu yol, çekirdek bazalisinde son bulur (Şekil P.Z, E). Güvercin ve yaban ördeğinde (Anas platyrhynos) , bu çekirdeğin hücreleri gaga, ağız boşluğu ve dilin dokunsal uyarımı üzerine boşaltılır (Witkovsky ve diğerleri, 1973; Berkhoudt ve diğerleri, 1981). Nukleus basalis'in boyutu, farklı sıralar arasında önemli ölçüde değişir ve gaga uzunluğu ve hassasiyetindeki karşılık gelen varyasyonları yansıtabilir. Güvercinin archistriatum'unu araştıran Zeier ve Karten (Zeier ve Karten, 1971), bazal çekirdeğin, neostriatumun üzerinde uzanan kısmına olduğu kadar, archistriatum'un ön çekirdeğine de yansıdığını bulmuşlardır. Archistriatum'un ön çekirdeği, ön serebral mesanenin dorsal kısmından gelişir (Şekil A.4, B). Yazarlar, işlevlerinin , bu baloncuğun ventral kısmından gelişen arkistriatumun diğer bileşenlerinden farklı olduğuna inanmaktadır (Bölüm 11.1.2 ve 11.1.3).
Madde 3.4. koku alma sistemi
Koku alma sisteminin birincil afferent lifleri koku alma ampulünde son bulur (Şekil P.1, A). Bu yapının boyutları çok değişkendir. Muhtemelen, bu varyasyonlar büyük ölçüde, bu türün makroosmat-mikroosmat serisi arasındaki konumunu yansıtmaktadır (Cobb, 1960a, b). Koku ampulünün çıkıntıları talamusta geçiş yapmadan ön beyne ulaşır . Güvercinlerde, koku alma sisteminden afferent bağlantıları alan beyin bölgeleri, koku alma lobunu (Şekil P.3 , A, B; 7) ve ventral hiperstriatum (Rieke ve Wenzel, 1978; Macadar ve diğerleri, 1980) içerir .
S.4. Beyin sapının orta kısmının projeksiyonları
Beyin sapı çeşitli sinir yapıları içerir. Beynin uzak kısımlarını, ayrıca beyin ve omuriliği birbirine bağlayan uzun yollar içerir . Kısa yükselen ve alçalan "birleştirici" lifler , beynin bir tarafındaki komşu yapıları birbirine bağlar ve komissural yollar, karşı taraflarını birbirine bağlar. Beyin sapı ayrıca kranial sinirlerin motor ve duyusal çekirdeklerini içerir. Burada, uzun süredir tanımlanmış olan, ancak bazıları Penfield (Penfield, 1938) ve J. ve M. Jeffersons tarafından keşfedilmiş olmasına rağmen, işlevleri hala çok sayıda çalışmanın konusu olan başka çekirdekler de vardır. (G. Jefferson, 1944; M. Jefferson, 1952) klinik çalışmalarda.
İnsanlarda, beyin sapının orta kısmındaki küçük bir doku alanına verilen hasar, bilinç kaybıyla ilişkilendirilir. Benzer bir koma, gövdenin medial bölgelerine zarar veren diğer memelilerde de bulundu (Lindsley ve diğerleri, 1950; French ve diğerleri, 1952). Bu merkezi sistemin işlevine ilişkin orijinal fikir , beynin genel durumunu kontrol etmesiydi. Diğer işlevlerin yanı sıra, bu sistemin uyanık-uyku döngüsünü modüle etmesi , aktivasyon ve uyanıklık seviyesini düzenlemesi ve dış sinyalleri beynin "yüksek" merkezlerine ulaşmamaları için filtrelemesi gerekiyordu . Daha yeni çalışmalar bu varsayımlardan bazılarını doğrulamıştır (Hobson ve Scheibel, 1980).
Memelilerde, beyin sapının bu çekirdeklerinden çıkan birkaç güçlü lif sistemi, orta beyin ve ön beynin hemen hemen tüm yapılarını innerve eder. Bu sistemlerden biri asetilkolinesteraz enzimini içerir, diğerleri monoaminler noradrenalin ve dopamin ile serotonin içerir. Bu nöron sistemlerinden bazıları şaşırtıcı özelliklere sahiptir. Örneğin norepinefrin içeren sistem böyledir. Onu oluşturan nöronların gövdeleri , mavi nokta adı verilen köprünün çekirdeğinde bulunur. Sıçanlarda, bu küçük hücre kümesinin aksonları beynin ve omuriliğin tüm ana bölgelerine uzanır (Ungerstedt, 1971). Bu tür fiber sistemlerin tümü çok yaygın olmasa da, bunların doğrudan etki alanları, kural olarak, belirli duyu yollarınınkinden çok daha geniştir.
Bazı durumlarda beyin sapından ön beyine doğrudan projeksiyonların varlığına dair kanıtlar dolaylı olsa da, kuş beyninde benzer sistemler mevcuttur. Bu bağlamda, monoamin içeren nöronların bağlantılarını belirlemek ve izlemek için kullanılan yöntemler hakkında birkaç söz söylenmelidir . Doku bölümleri formaldehit buharı (Falck ve diğerleri, 1962) veya glioksilik asit (Björklund ve diğerleri, 1975) ile işlenir . Bu maddelerle reaksiyona giren monoaminler, yoğun flüoresan ürünler oluşturur; monoamin içeren hücreler daha sonra ultraviyole ışıkta bir mikroskop altında gözlemlenebilir. Serotonin içeren hücreler sarı bir parıltıya sahipken , norepinefrin ve dopamin monoaminlerini içeren hücreler sarı-yeşil bir parıltıya sahiptir. Bu iki monoaminin farklılaşması ek , genellikle farmakolojik yöntemler gerektirir.
ve muhabbet kuşunun (Melopsittacus undulatus) (Shiosaka ve diğerleri , 1981) beyin sapında monoamin içeren sinir hücresi kümeleri tanımlanmıştır . ) . Orta beynin tegmentumunda ve ayrıca pons ve medulla oblongata'da, özellikle mavi nokta alanında katekolamin içeren nöronlar bulundu (Şekil A.7). Memelilerde , orta beynin tegmentumundaki hücre grupları dopamin içerirken, daha kaudal kümeler, özellikle locus coeruleus ve bununla ilişkili bölgeler, norepinefrin içerir (Lindvall ve Björklund, 1978). Bu hücre gruplarının hem memelilerde hem de kuşlarda farklı ön serebral hedefleri vardır. Memelilerde, bir grup dopamin içeren nöronlar (substantia nigra hücreleri) kaudat çekirdeğe ve pu -topa yansıtılır (Lindvall ve Björklund, 1978). Bu oluşumlar serebral hemisferlerin içinde yer alır ve hareketleri kontrol eden karmaşık bir çekirdek grubunun parçasını oluşturur. Kuşlarda, katekolamin içeren orta beyin tegmental hücrelerinin hedef bölgelerinden biri yükseltilmiş paleostriatumdur (Brauth ve diğerleri, 1978; Kiff ve Brauth, 1986). Bu bağlamda, bir yandan kaudat çekirdeğin ve putamenin benzer bir embriyolojik kökeninin ve diğer yandan birincil beyin mesanesinin ventral bileşeninden yükseltilmiş paleostriatumun gerçeği (Şekil P.4, A) , B) ilgi çekicidir . Memelilerde başka bir dopamin içeren hücre grubunun adresleri koku alma tüberkülü ve akkumbens'tir (Şekil A.7). Zebra ispinozlarında (Peophila guttata), benzer bir katekolaminerjik hücre grubu, koku alma lobunu ve muhtemelen n. akumbens'i innerve eder (Lewis ve diğerleri, 1981). Karşılaştırmalı nöroanatomi verileri , kuş orta beyninin tegmentumundaki katekolaminerjik hücrelerin dopamin içerdiğini göstermektedir. Ancak bu yalnızca bir varsayımdır. Bu kanıtlanmış olsa bile, dopaminerjik innervasyonun Şekil 1'de solda gösterilen bazal ön beyin yapılarıyla sınırlı olmadığı vurgulanmalıdır . S.7. Davis ve diğerleri (1983), dopaminin Wulst ve civciv ventral hiperstriatumunda da bulunduğunu göstermiştir (ayrıca bkz. Kitt ve Brauth, 1986).
Daha önce belirtildiği gibi, locus coeruleus'tan gelen sinir lifleri sistemi, memeli ön beyninde yaygın olarak dağılmıştır. Muhabbet kuşlarında, önbeynin tüm ana bölümlerinde katekolamin içeren sinir lifleri bulundu. Bu liflerin bazıları, belki de tümü, mavi noktadan ve komşu çekirdeklerden geliyor gibi görünmektedir (Tohyama ve diğerleri, 1974; Takatsuki ve diğerleri, 1981). Medulla oblongata'da bulunan
katekolamin içeren nöronların projeksiyonları hakkında çok az şey bilinmektedir .
Resim: S.7. Beyin sapındaki monoamin içeren nöronların olası ön beyin projeksiyonları. Solda, orta beyin çatısında katekolamin içeren hücre gövdelerinin lateral ve medial birikimleri gösterilmektedir. Tavuklar, güvercinler, papağanlar ve zebra ispinozları üzerinde yapılan çalışmalarda elde edilen veriler, bu hücrelerin aksonlarının yükseltilmiş paleostriatuma (9), koku alma lobuna (10) ve akumbenslere ( 11) çıkıntı yaptığını göstermektedir. Memelilerin beynindeki karşılık gelen projeksiyonlara benzetilerek , bu hücrelerin vücutlarında bulunan katekolamin'in dopamin olduğu varsayılabilir. Sağda, mavi noktanın içinde ve çevresinde hücre kümeleri (12) gösterilmektedir. Bu hücre gruplarını içeren alan, diyagramın sol tarafında gösterilenin kaudalinde yer alır. Muhabbet kuşunda, variköz katekolamin içeren aksonlar ön beynin tüm ana bölümlerine uzanır; muhtemelen kaynakları, norepinefrin içeren mavi nokta hücrelerinin birikmesidir. Ön beyindeki liflerin dağılımı, birkaç kuş türü üzerinde yapılan çalışmada ortaya çıktı; Beyin sapındaki katekolamin içeren nöronların organizasyonu, tavuk beyni üzerinde yapılan çalışmalara dayanmaktadır (Dube ve Parent, 1981). Teraziye uyulmuyor. Pontin tegmentumun 1 - ventral ve 2 - dorsolateral hücre grupları; 3 - görsel çatı ; 4 - ventral hiperstriatum; 5 - dorsal hiperstriatum; 6 - ek hiperstriatum; 7 - ektotriyum; 8 - neostriatum. Daha ayrıntılı açıklamalar için metne bakın.
Serotonin içeren hücre gövdeleri , tavukların orta beyninin kaudal kısmında ve medulla oblongata'da bulunur (Dube ve Parent, 1981). Bu hücrelerin artan projeksiyonları henüz incelenmemiştir .
Güvercin ön beyni asetilkolin içerir (Aprison ve Takahashi, 1965), ancak kesin lokalizasyon verileri dolaylıdır ve esas olarak asetilkolinesteraz enziminin histokimyasal olarak tespit edilebilir dağılımının çalışmasına dayanmaktadır . Bununla birlikte, nöronlardaki asetilkolinesteraz içeriğinin , bu nöronların kolinerjik doğasının yeterli kanıtı olmadığını hesaba katmak önemlidir . Asetilkolinesteraz tavuk (McCabe ve ark., 1982) ve güvercin (Vischer ve ark., 1982) beyninin birçok bölgesinde bulunur. Tavukların ön beyinlerinde asetilkolinesteraz için lekelenme dağılımının mikroskobik bir resmi, bu enzimin bir kısmının sinir liflerinin terminallerinde bulunabileceğini göstermektedir (McCabe ve diğerleri, 1982). Asetilkolin analoglarını bağlayan reseptörler kuşların ön beyinlerinde de bulunur ve konsantrasyonları bölgeye göre değişir (Bradley ve Noth, 1981; Jerusalinsky ve diğerleri, 1981; Vischer ve diğerleri, 1982). Bu reseptörlerin varlığı, aynı beyin bölgesindeki sinir terminallerindeki asetilkolinesteraz içeriğine ilişkin verilerle birlikte, bu bölgeyi innerve eden afferent liflerin kökeni sorusunu gündeme getirmeyi mümkün kılar. Tavuklarda, bu liflerin bir kısmı ön beynin iç sistemine aittir: bir bölgenin hücre gövdeleri diğer bölgelere yansıtılır (Davies ve Horn, 1983). Ancak bazı memelilerde olduğu gibi ön beyindeki kolinerjik nöronların iç sistemine ait olup olmadığı bilinmemektedir . İkincisinde, nükleus bazalis, komşu septal çekirdekler ve diyagonal demetin çekirdeği, kolinerjik afferentlerin ana kaynaklarıdır (Lewis ve Shute, 1967; Kimura ve diğerleri, 1981; Mesulam ve diğerleri, 1983). ). Tavuklarda asetilkolinesteraz içeren septal bölge hücreleri ön beyin üzerine yansıtılır (Davis ve Horn, 1983). Tavukların nükleus bazalis (Şekil P.3, A) hücre gövdeleri de bu enzimden büyük miktarda içerir (McCabe ve diğerleri, 1982), ancak bu hücrelerin projeksiyonları henüz çalışılmamıştır. Bununla birlikte, kuşların ve memelilerin bazal gangliyonları arasında bir yazışma olduğuna dair net bir kanıt yoktur; karşılaştırmanın zorlukları, bu çekirdeğin anatomisindeki çarpıcı tür farklılıklarının varlığıyla daha da karmaşık hale gelir (Gorry, 1963). Memelilerde asetilkolinesteraz içeren bazı sinir lifleri orta beynin tegmentumundan gelir; kortikal bölgeleri innerve ettiklerine inanılmaktadır (Shute ve Lewis, 1967). Kuşlarda benzer bir sistemin varlığına dair veriler henüz mevcut değildir.
S.5. Serebral hemisferlerin inen yolları
Decerebrate kuşlar, yani ön beyin yarıküreleri çıkarılmış kuşlar nispeten hareketsizdir. Ancak, beyinsiz civcivlerin dürtüldüğünde yürüyebildikleri ve havaya fırlatıldıklarında uçabildikleri uzun zamandır bilinmektedir (Flourens, 1824, aktaran Liddell, 1960). Beyinsiz yaban ördeği görünüşte normal birçok hareket yapar ve örneğin tüylerini fırçalayabilir, başlarını kaşıyabilir, kanatlarını çırpabilir ve kuyruklarını oynatabilir (Phillips, 1964). Sonuç olarak, bu hareketleri sağlayan ana sinir mekanizmaları daha düşük seviyelerde lokalizedir. Benzer bir durum kedilerde de görülmektedir. Bu hayvanda yürüme sırasında arka bacakların ritmik hareketlerini sağlayan mekanizma omurilikte yer almaktadır (Shik ve Orlovsky, 1976). Normal bir kedide bu mekanizmalar, beyin sapına ve omuriliğe giden uzun inen yollardan hareket ederek daha yüksek beyin bölgelerinin kontrolü altındadır. Kuşların , beyin sapı ve omuriliğin nöral mekanizmalarını düzenleyen ön beyinden aşağı inen benzer yolları var mı? Anatomik kanıtlar, işlevleri henüz tam olarak anlaşılmamış olsa da, bu tür yolların varlığını açıkça göstermektedir. Bir alçalan yol , birkaç çekirdekten oluşan karmaşık bir yapı olan arşistriatumun (Şekil P.Z, D) bir bölümünde başlar (Zeier, Karten, 1971). Bu çekirdekler, bağlantıları ve olası embriyonik kökenlerine göre iki ana gruba ayrılır (Şekil A.4, B). Posterior ve medial bileşenler hipotalamus ile ilişkilidir ve bu nedenle vissero-endokrin fonksiyonlarda yer alabilir (Zeier ve Karten, 1971). Archistriatum'un ön kısmı ( dorsal bileşeni ile birlikte ön ve orta archistriatum ) somatik, sensorimotor sistemle ilişkilidir. Ön beyin hemisferlerinden gelen bu inen yollar , archistriatum'un bu bileşeninde başlar . Bu yolun terminal alanı, talamusu, optik çatıyı ve beyin sapının merkezi çekirdekleri dahil olmak üzere beyin sapının orta ve arka kısımlarının birkaç çekirdeğini içerir. İnen yollar da omuriliğin ön kısmına geçer. Görünüşe göre, bu yollar hareketlerin düzenlenmesinde yer alan subtelensefalik yapıları etkileyebilir . Uydukları bir dizi yapı duyusal bilginin iletilmesinde yer aldığından ( örneğin , medulla oblongata'nın görsel çatısı ve somatosensoriyel çekirdekleri) duyusal sinyallerin akışını da düzenleyebilirler. Ek olarak, arşistriatumun sensorimotor bölümü , mavi nokta alanında oldukça geniş terminal dallarına sahip oldukları için davranışın daha genel yönlerini etkileyebilir .
Ön beynin ötesine uzanan bir başka sinir lifi kaynağı da Wulst'tur. Bu tahminler, kuzgun (Corvus corax), pembe yanaklı cennet papağanı (Agapornis roseicollis) (Adamo, 1967) ve mağara baykuşunun (Speotyto cypicilaria ) yanı sıra evcil tavuklarda (Adamo, 1967; Bradley ve diğerleri, 1985) incelenmiştir. ) \( Karten ve diğerleri, 1973). Özellikle iyi gelişmiş bir Wulst'a sahip olan baykuşun inen iki yolu vardır. Bunlardan biri Wulst'un küçük ön bölümünden çıkar ve beyin sapının çekirdeklerinde, özellikle kırmızı çekirdekte ve arka beynin medial bölgesinin retiküler oluşumlarında son bulur (Karten, 1971). Diğer omurgalı sınıflarında da bulunan bu çekirdeklerin, lokomotor aktivitenin çeşitli yönlerini düzenleyerek doğrudan veya omuriliğe geçtikten sonra çıkıntı yaptığına dair kanıtlar vardır (bkz . Pearson ve Pearson, 1976; Brodal, 1981). İnen bazı ön Wulst lifleri omuriliğe kadar uzanır . Güvercindeki ön Wulst'un somatosensoriyel sinyaller aldığını hatırlamak ilginçtir : duyusal sinirlerin doğrudan elektriksel stimülasyonu için uyarılmış potansiyeller bu alanda kaydedilebilir (Delius ve Bennetto, 1972). Baykuşta benzer bir duyusal alanın var olup olmadığı bilinmemektedir . Eğer öyleyse, ön Wulst'un organizasyonu bazı memelilerdeki motor korteksinkine benzer : motor korteks hücreleri, derinin ve derin dokuların uyarılması üzerine uyarılabilir (Brooks ve Stoney, 1971; Asanuma, 1975).
Wulst'un geniş arka kısmı görsel bir işleve sahiptir (bölüm A.3.1). Wulst'un bu bölümünden , sinir lifleri beynin çeşitli bölümlerine gider, ancak esas olarak görsel yolun aktarma bağlantılarına gider. Bu bağlantılardan bazıları ön beyin içinde yer alır, örneğin periectostriatal kuşak ve arşistriatumun dorsal kısmı (bkz. Bölüm A.3.1); talamusun ana optik çekirdeği ve optik çatı gibi diğerleri telensefalonun altındadır. Archistriatum'un sensorimotor kısmından karşılık gelen projeksiyonlar gibi bu projeksiyonlar, duyusal sinyallerin merkezi sinir sistemine akışını düzenler. Görsel dürtü durumunda, bu düzenleme, görsel sistemin kendisinin bağlantıları ve görsel-motor koordinasyonuna katılabilecek alanlar aracılığıyla gerçekleştiriliyor gibi görünüyor .
S.6. Özet
Kuş beyninin birkaç ayrı duyusal yansıtma alanı vardır. Görsel, işitsel ve somatosensoriyel sistemlerde bilgi alan en az iki ayrı alan vardır . Orta beynin belirli alanlarından, hareket ve duyusal girdilerin düzenlenmesinin gerçekleştirilebileceği yollar ayrılır. Beyin sapındaki hücre grupları doğrudan ön beyin üzerine projekte olur ve ön beyin fonksiyonlarının yerel ve genel düzenlemelerini gerçekleştirebilir.
140
EDEBİYAT
Adamo NJ Tavuk, kuzgun ve Afrika muhabbet kuşundaki hiperstriatal bölgelerden gelen efferent liflerin bağlantıları, J. comp. Neurol., 1967, 131, 337-356.
Aprison MH, Takahashi R. Kuş merkezi sinir sisteminin biyokimyası.
II. Güvercin beyninin birkaç ayrı alanında 5-Hidroksitriptamin, asetilkolin, 3,4-dihidroksifenil-etilamin ve norepinefrin, J. Neurochem., 1965, 12, 221-230.
Asanuma H. Motor korteks, Physiol içindeki nöronların sütunlu düzenine ilişkin çalışmadaki son gelişmeler. Rev., 1975, B5 143-156.
Benowitz L. Kuş telensefalonunun fonksiyonel organizasyonu. İçinde: Telensefalonun karşılaştırmalı nörolojisi (ed. S. O. E. Ebbesson), 1980, s. 389-421, Plenum Yayıncılık, New York.
Berkhoudt H., Dubbelman JL, Zeilstra S. Yaban ördeği Anas platyrhynchos L. IV'teki trigeminal sistemin somatotopisi üzerine çalışmalar . Nukleus basalis'te dokunsal temsil, J. comp. Neurol., 1981, 196, 407-420.
Björklund A., Falck B., Lindvall O. Formaldehit veya glioksilik asit yoğunlaşmasından sonra hücresel monoaminlerin mikrospektraflorometrik analizi. İçinde: Beyin araştırmalarında yöntemler (ed. PB Bradley), 1975, s. 249-294, Wiley, New York.
Bonke B. A., Bonke D., Scheich H. Gine tavuğundaki (Numida meleagris) işitsel ön beyin çekirdeklerinin bağlantısı , Celi Tissue Res., 1979, 200, 101-121.
Güvercin , J. comp. Neurol., 1968, 133, 523-541.
Board RL Kuşların işitme sisteminin anatomisi, Ann. NY Acad. Sci., 1969, 167, 186-198.
Boord RL, Rasmussen GL Güvercinin koklear çekirdeği üzerindeki koklear ve lagenar sinirlerin projeksiyonu, J. comp. Neurol., 1963, 120, 463-475.
Bradley P., Horn G. Civciv beyninde hiperstriatum ventrale'nin afferent bağlantıları, J. Physiol., Londra., 1978, 278, 46P.
Bradley P., Horn G. Baskı. Civciv beyninin bazı bölümlerindeki kolinerjik reseptör bölgelerine ilişkin bir çalışma, Exp. Brain Res., 1981, 41, 121-123.
Bradley P., Davies D., Horn G. Evcil civcivde (Gallus domesticus) hyperstriatum ventrale'nin bağlantıları, J. Anat., 1985, 140, 577-589.
Brauth SE, Ferguson JL, Kitt CA Güvercin paleostriatumuyla ilgili prosensefalik yollar (Columba livia), Brain Res., 1978, 147, 205-221.
Brodal A. Klinik tıpla ilgili olarak nörolojik anatomi (3. baskı) 1981, Oxford University Press, New York.
Brooks VB, Stoney SD Jr. Motor mekanizmalar: piramidal sistemin motor kontroldeki rolü, A. Rev. Physiol., 1971, 33, 337-392.
Brown MW, Horn G. Piliç beynindeki nöronal plastisite: görsel deneyimin hiperstriatal nöronlar üzerindeki elektrofizyolojik etkisi, Brain Res., 1979, 162, 142-147.
Cobb S. Kuşların koku soğancığının boyutu üzerine bir not, Epilepsia, Perspect. Biol. Med., 1960b, 3, 383-408.
Cobb S. Kuş beyninin karşılaştırmalı anatomisi üzerine gözlemler, Perspect. Biol. Med., 1960b, 3, 383-408.
Cohen DH, Karten HJ Kuş beyninin yapısal organizasyonu: Genel Bakış. İçinde: Kuşlar, beyin ve davranış (ed. IJ Goodman ve M.W. Schein), 1974, s. 29-73, Academic Press, New York.
Cowan WM, Adamson L., Powel TPS Avlan görsel sisteminin deneysel bir çalışması, J. Anat., 1961, 95, 545-563.
Davies DC, Horn G. Piliç hyperstriatum ventrale'nin varsayılan kolinerjik afferentleri: kombine bir asetilkolinesteraz ve retrograd floresan etiketleme çalışması, Neurosci. Lett, 1983, 38, 103-107.
Davies DC, Horn G., McCabe BJ Evcil civcivlerde telensefalik katekolamin seviyelerindeki değişiklikler. Yaş ve görsel deneyimin etkileri. Devi. Brain Res., P98Ö, 10, 2'51-2'55.
Delius JD, Bennetto K. Kuşların ön beynine kutanöz duyusal projeksiyonlar, Brain Res., 1972, 37, 205-221.
Delius JD, Runge TE, Oeckinghaus H. Güvercin ön telensefalonuna kısa gecikmeli işitsel projeksiyonlar, Exp. Nörol." 1979, 63, 594-609.
Dubö L., Ebeveyn A. Kuş beynindeki monoamin içeren nöronlar:
Flüoresan ve asetilkolinesteraz histokimyası yoluyla tavuğun (Gallus domesticus) beyin sapının incelenmesi , J. comp. Neurol., 1981" 196, 695-708.
Erulkar SD Güvercin beynindeki dokunsal ve işitsel alanlar, J. komp. Neurol., 1955, 103, 421-457.
Felix B., Kesar S., Roesch T. Landes kazında somatik uyarılmış yanıtların merkezi lokalizasyonu, Exp. Brain Res., 1983, BZ, 173-182.
French JD, Amerongen FK von, Magoun HW Archs Neurol maymununun beyin sapında aktive edici bir sistem. Psychiat., Chicago, 1952, 68, 577-590.
Falck B., Hillarp N.-A., Thiene G. f Torp A. Formaldehit ile yoğunlaştırılmış katekolaminlerin ve ilgili bileşiklerin floresansı, J. Histochem. Cytochem., 1962, 10, 348-354.
Fuxe K., Ljunggren L. Güvercin üst beyin sapındaki monoaminlerin hücresel lokalizasyonu, J. comp. Neurol., 1965, 125, 355-382.
Gorry JD Meynert'in ganglion bazalesinin karşılaştırmalı anatomisi üzerine çalışmalar, Açta anat., 1963, 55, 51-104.
Harman AL, Phillips RE Tıklama uyaranlarıyla uyarılan kuş orta beyni, talamus ve ön beyindeki yanıtlar, Exp. Neurol., 1967, 18, 276-286.
Hobson JA, Scheibel AB Beyin sapı çekirdeği: sensorimotor entegrasyon ve davranışsal durum kontrolü, Neurosci. Res. Program Bülteni, 1980, 18, 1-173.
Hunt S.P. , Webster K.E. Güvercinlerde Thalamo-hiperstriat karşılıklı ilişkileri, Brain Res., 1972, 44, 647-651.
Ikeda H., Gotoh J. Tavuğun merkezi sinir sisteminde monoamin içeren hücrelerin dağılımı, Jap. J. Pharmac., 1971, 21, 763-784.
Ilyichev V.D., Gurin S.S., Temchin A.N. , Voronetsky V.S. Bir güvercinin işitme sisteminin biyolojik sinyali ve fonksiyonel özellikleri, Zhurn. genel biol., 1970, 31, 268-274.
Jefferson G. Bilincin doğası, Br. tıp J., 1944, 1, 1-15.
Jefferson M. Beyin sapı lezyonlarıyla ilişkili değişen bilinç, Brain, 1952, 75, 55-67.
Jerusalinsky D., Aguilar JS, Brusco A., De Robertis E. Civciv beyninin çeşitli alanlarında muskarinik reseptörlerin ve asetilkolinesteraz aktivitesinin ontogenezi,
J._ _ Neurochem., 1981, 37, 1517-1522.
Kaien B. Civciv telensefalonunda beyin çekirdeklerinin embriyogenezi, Ergebn. Anat. EntwGesch., 1962, 36, 62-82.
Kappers C. VA, Huber GC, Crosby EC İnsan da dahil olmak üzere omurgalıların sinir sisteminin karşılaştırmalı anatomisi, 1936, MacMIIIan, New York.
Karten HJ Güvercinlerde (Columba livia) yükselen işitsel yolun organizasyonu. I. Alt kollikulusun diensefalik projeksiyonları (nucleus mesensefali lateralis, pars dorsalis), Brain Res., 1967, 6, 409-427.
Karten HJ Güvercinlerde (Columba livia) yükselen işitsel yol. II. Nucleus ovoidalis thalami'nin telensefalik projeksiyonları, Brain Res., 1968, 11, 134-153.
Karten HJ Kuş telensefalonunun organizasyonu ve amniyotik telensefalonun filogenisi üzerine bazı spekülasyonlar, Ann. NY Acad. Sci., 1969, 167, 164-179.
Karten HJ Effereht projeksiyonları, Anat Rec., 1971, 169, 353.
Karten HJ, Hodos W. Güvercindeki rotundus çekirdeğinin telensefalik projeksiyonları (Columba livia) , J. comp. Neurol., 1970. 140, 35-52.
22*
Karten HJ, Hodos W., Nauta W 7 . JH t Revzin AM Asya telensefalonunun “görsel Wulst”unun sinirsel bağlantıları. Güvercin (Columba livia) ve baykuşta (Speotyto cunicularia) deneysel çalışmalar, J. comp. Neurol., 1973, 150, 253-278.
Kimura H., McGeer PL, Peng JH, McGeer EG Kedide kolin asetiltransferaz immünohistokimyası ile incelenen Merkezi kolinerjik Sistem, J. comp. Neurol., 1981, 200, 151-201.
Kirsch M., Coles RB, Leppelsack HJ Uyanık sığırcığın (Sturnus vulgaris) frontal neostriatumundaki yeni bir işitme alanından kayıt yapan birim , Exp. Yağmurda Res., 1980, 38, 375-380.
Kitt CA, Brauth SE Güvercin orta beyin ve isthmal hücre gruplarından telensefalik projeksiyonlar. II. Nigral kompleksi, J. comp. Neurol., 1986, 247,
110.
Knudsen E. L, Konishi M. Uzay ve frekans, baykuşun işitsel orta beyninde ayrı ayrı temsil edilir, J. Neurophysiol., 1978, 41, 870-884.
Konishi M„ Knudsen EI Sinir işitsel alan teorisi: baykuşta işitsel temsil ve önemi. İçinde: Çoklu kortikül alanları (ed. C. Woolsey), 1983, Humann Press, Clifton, NJ
Kuhlenbeck H. Civcivdeki korteks telensefalinin ontogenetik gelişimi ve filogenetik önemi, J. comp. Neurol., 1938, 69, 273-301.
Lepelsack HJ, Vogt M. Ötücü bir kuşun ön beynindeki işitsel nöronların türe özgü seslerle uyarılmaya tepkileri, J. comp. Physiol., 1976, A. 107, 263-274.
Lewis JW, Ryan S. M, Arnold A. P., Butcher LL Zebra ispinozunda X alanına katekolaminerjik projeksiyon için kanıt, J. comp. Neurol., 1981, 196, 347-354.
Lewis PR, Shute CCD Kolinerjik limbik sistem: hipokampal formasyona, medial kortekse, yükselen retiküler sistemin çekirdeklerine ve subfornikal organa ve supra-optik tepeye projeksiyonlar, Brain, 1967, 90, 521-540.
Liddell EGT Reflekslerin keşfi, 1960, Clarendon Press, Oxford.
Lillie FR Civcivlerin gelişimi, 1980, Henry Holt, New York.
Lindsley DB, Schreiner LH, Knowles WB, Magoun HW Kedi Electroenceph'te chronie beyin sapı lezyonlarını takiben davranışsal ve EEG değişiklikleri. klinik Neurophysiol., 1950, 2, 483-498.
Lindvall O., Björklund A. Sıçan Merkezi sinir sisteminde katekolamin nöronlarının organizasyonu. İçinde: Psikofarmakoloji El Kitabı (eds. LL iversen , SD iversen ve SH Snyder), 1978, Cilt. 9, s. 139-231, Plenum Yayıncılık, New York.
Macadar AW, Rausch LJ, Wenzel BM, Hutchison LV Güvercinlerdeki koku alma yolunun elektrofizyolojisi, J. comp. Physiol., A. 1980, 137, 39-46.
McCabe BJ, Horn G., McGrath G. Civciv telensefalonunda asetilkolinesterazın dağılımı, J. Anat., 1982, 134, 600-601.
Macphail E. M. Omurgalılarda beyin ve zeka, 1982, Clarendon Press, Oxford.
.Mesulam MM, Elliot JF, Levey A. L, Wainer B. H. Bazal ön beyin tarafından korteksin kolinerjik innervasyonu: al yanaklı maymunda septal alanın sitokimyası ve kortikal bağlantıları, diyagonal bant çekirdekleri, nükleus bazalis (substantia innominata) ve hipotalamus, J. komp. Neurol., 1983, 214, 170-197.
Nauta WJH, Karten HJ Serebra 1 korteksinin ataları üzerinde yan ışıklarla omurgalı beyninin genel bir profili. İçinde: Nörobilimler, ikinci çalışma programı (ed. F.O. Schmitt), 1970, s. 7-26. Rockefeller University Press, New York.
(DDM'de Lear'da, Gerfen C. R „ Cowan WM Civcivde ipsilateral retinofugal projeksiyonun gelişimi ve kısıtlanması. Devi. Brain Res., 1983, 10,
109.
Parker DM, Delius JD Güvercinin ön beyin bölgesindeki görsel uyarılmış potansiyeller, Exp. Brain Res., 1972, 14, 198-209.
Pearson R. Kuş beyni. Academic Press, Londra, 1972.
Pearson R., Pearson L, Omurgalı beyni, Academic Press, Londra, 1976.
Penfield W. İnsanın serebral korteksi. 1. Serebral korteks ve bilinç. Arch Neurol. Psychiat., Chicago, 1938, 40, 417-442.
Pettigrew JD, Ronishi M. Peçeli baykuşun (Tyto alba) görsel Wulst'unda yönlendirme ve binoküler eşitsizlik için seçici nöronlar, Science, 1976, 193, 675-678.
Phillips RE Yaban ördeğindeki vahşilik: beyin lezyonlarının ve uyarımın “kaçış davranışı” ve üreme üzerindeki etkileri, J. comp. Neurol., 1964, 122, 139-155.
Rieke GK, Wenzel BM Güvercin koku ampulünün ön beyin projeksiyonları, J. Morph., 1978, 158, 41-56.
Ritchie TC, Cohen DH Kuş tektofugal görsel yolu: telensefalik hedefinin projeksiyonları, ekstostriatal kompleks, Proc. sos. Neurosci., 1977, 3, 94.
Sachs MB, Sinnott JM Redwinged Blackbird'ün (Agelaius phoenicus) nükleus magnocellularis ve nükleus angularisindeki tek hücrelerin tonlarına yanıtlar . J komp. Physiol., 1978, A126, 347-361.
Scheich H. Civcivlerin işitsel neostriatumundaki iki sütunlu sistem: 2-deoksiglikozdan kanıtlar, Exp. Brain Res., 1983, 51, 199-205.
Scheich H., Bonke BA, Langner G. Gine tavuğundaki bazı işitsel çekirdeklerin 2-deoksiglikoz tekniği, Cell ile gösterilen fonksiyonel organizasyonu. Doku Res., 1979a, 204, 17-27.
Scheich H., Langner G., Bonke D. Gine tavuğunun (Numida meleagris) işitsel neo striatumundaki birimlerin türe özgü çağrılara ve sentetik uyaranlara duyarlılığı. II: iambus benzeri aramaların ayrımı, J. comp. Physiol., 1979b, A132, 257-276.
Shik ML, Orlovsky GN Lokomotor otomatizminin nörofizyolojisi, Physiol. Rev., 1976, 56, 465-501.
Shiosaka S., Takatsuki K., Inagaki S., Sakanaka M., Takagi H., Senba E., Matsuzaki T., Tohyama M. Topografik atlas of somatostatin-containing neuron system in the avianbeyin ile ilgili olarak katekolamin içeren nöron Sistem. II. Mesencephalon, eşkenar dörtgen ve omurilik, J. comp. Neurol., 1981, 202, 115-124.
Shute CC, Lewis PR Yükselen kolinerjik retiküler sistem: neokortikal, koku alma ve subkortikal projeksiyonlar, Brain, 1967, 90, 497-520.
Stingell W7._ _ _ Verleichend morfologische untersuchungen am Vordenhirn der Vogel auf sitologischer ve celloarchiektonischer Grundlage. 1958, Hebling ve Lichtenhahn, Basle.
stoppe.pe Whitfield/. C. Güvercindeki beyin sapı çekirdeklerinden birim yanıtları, J. Physiol., 1961, 158, 165-177.
Takatsuki K., Shiosaka S., Inagaki S., Sakanaka M., Tagaki H., Senba E., Matsuzaki T., Tohyama M. Topografik atlas of somatostatin-containing neuron system in the avianbeyin, ilişkin olarak katekolamin içeren nöron sistemi. I. Telencephalon ve diensefalon, J. comp. Neurol., 1981, 202, 103-113.
Tohyama M., Maeda T., Hashimoto J., Shrestha GR, Tamura O., Shimizu N. Locus coeruleus'un karşılaştırmalı anatomisi. I. Kuşun arka bölgesindeki katekolamin içeren nöronların organizasyonu ve artan projeksiyonları, Melopsittacus undulatus, J. Hirnforsch., 1974, 15, 319-330.
Sıçan beynindeki monoamin yollarının Vngerstedt U. Stereotaksik haritalaması. Acta fizyol. taramak suppi. 1971, 367, 1-48?
Vischer A., Cuânod M., Henke H. Nörotransmitter reseptör ligand bağlanması ve güvercin görsel sisteminde enzim bölgesel dağılımı, J. Neurochem., 1982, 38, 1372-1382.
Wallenberg A. Der Ursprung des Tractus isthmo-striatus (öder bulbostriatus) der Taube, Neurol. Zent., 1903, 22, 98-101.
Wilson P. Evcil civcivlerde görsel hiperstriatumun organizasyonu. II. Tek birimlerin alıcı alan özellikleri, Brain Res., 1980, 188, 333-345.
Witkovsky P., Zeigler HP, Silver RA Güvercindeki çekirdek bazalis'in tek birim analizi, J. comp. Neurol., 1973, 147, 119-128.
Zaretsky MD, Ronishi M. Avian telencephalon'daki Tonotopik organizasyon, Brain Res., 1976, 111, 167-171.
Zeler H., Karten HJ Güvercin arşivi: afferent ve efferent bağlantıların organizasyonu, Brains Res., 1971, 31, 313-326.
KONU DİZİNİ
İtalik olarak yazılan rakamlar şekillere atıfta bulunmaktadır.
Adrenokortikotropik hormon (ACTH) 63, 203
232, 234'ün aktif bölgesi
Axodendritic dikenler 235
Axospinous sinapslar 77, 235-254
Amigdala 282, 283
memelilerde 275
Archistriatum'un amigdala kısmı
282
Amino asitler 50, 261
Hafıza kaybı 110-116, 135-138, 283, 290-297
diensefalik 136, 295
Kuşlarda androjenler ve şarkı 219
Aplysia (Aplysia) 25, 26, 30, 32, 250, 297
Archicortex (eski ağaç kabuğu) 158, 281
Archistriatum 91, 119, 120, 315,
316, 317, 328
arka çekirdekler 273, 328
ve PMVG 272, 276
ve davranış 273-275
medial çekirdekler 273, 274, 317
ön çekirdekler 317, 323, 328
ara 264, 272, 273,
328
İlişkilendirmeli öğrenme 38, 126-147, 275, 291
• ve tanıma 135-138
PMVG 126-135, 143-147, 171
Duygusal durum (duygular) 203, 273-275
Asetilkolin 253, 277, 327
Asetilkolinesteraz 278-279, 324, 327
Bazal çekirdek 270, 316, 323, 327
Sincap (dönen) çark 67, 68, 186-189, 220
Karın derisi refleksi 293
PMVG 155-159'da izlerin arabellekte saklanması
Superior colliculus 17, 265, 266 Olgun doğan türler 39, 40, 41
olgunlaşmamış 39, 40 Işık ve dendritik maruziyet
dikenler 77, 242
yatkınlık 182-189, 195, 306
Uyarıcı postsinaptik potansiyel (EPSP) 25-29, 30, 31, 242, 247-248
Fiber muhabbet kuşları (Melopsittacus undulatus) 325, 326
Kuzgun (Corvus corax) 328
Gri karga 266 (Corvus corone)
Geçici Depolama (X') 155-161, 168, 169, 296
Vulst 55, 98, 302, 313, 315
hasar 119, 178
çift taraflı 108-109, SW, 113, 114, 193-194
tek taraflı 116-118, 152, 153
iletişim 262, 317, 319, 320, 322, 328-329
katekolamin seviyesi 205, 207, 213, 325
6-Hidroksidopamin (6-ODA) 204, 205, 212
5-hidroksitriptamin (serotonin) 32, 277, 324, 326
Hiperstriatum ventral 235, 315, 316, 317, 323, 325
görsel yollar 263, 266, 320
ve alan L 95, 269, 321
ötücü kuşların kuyruk kısmı 161
hasar 113, 122
radyo imza 91, 93, 94, 95, 97, 143
asetilkolinesteraz 278 içeriği
isteğe bağlı 228, 229, 238, 240, 315, 316, 317
radyo imzası 91, 98
iletişim 262, 276, 319
elektrik stimülasyonu 200-201
sırt 91, 92, 113, 276, 277,
316, 317
3 H-lisin 55 emilimi
ara 91, 113, 276, 315,
322
Hipotalamus 274, 275, 282, 283, 284, 313, 314, 328
Hipokampus (hipokampal bölge) 21, 121, 278, 279, 281, 283, 315, 316, 317
hafızadaki rol 135, 158, 275, 284
sinaptik değişiklikler 242, 253
Ana duyu çekirdeği 270 L-glutamat 213
Glikoz 96. Ayrıca bkz. 2-deoksiglukoz
Güvercinler 65-67, 272, 273, 283
paleostriatal kompleks 165, 273
duyusal yollar 262, 270, 318-323
Hormonlar 63, 254
Kaz grisi 263, 270, 320, 322
Motor aktivite, düzenleme 212, 270, 327-328 2-deoksiglukoz 96-98, 143, 269 Deserebrasyon 118, 327 Diasilgliserol 253 Göz hakimiyeti 204, 213, 303 Dorsal supraoptik komissür 65-71, 319
Dopamin 203, 206-208, 277, 324, 325
DSP-4 (N-(2-kloroetil)-N -etil-bromo-benzil aminohidroklorür) 208-
212, 213, 219, 222
Arka beyin 272, 314
Oksipito-mezensefalik yol 271, 272, 273
Zebra ispinozları (Peophila gutta ta) 219, 325
Görsel korteks 204, 242, 264, 301 - 305
çatı, dahil etme 8 H-lizin 55-56
ve bağımlılık yapan 17, 18, 20
inen yollar 271-272, 273 beynin diğer bölümlerine giden yollar 318-320
PMVP 264 ile iletişim
stimülasyon 41, 65-71, 139, 174 Optik sinir 202, 313, 318
uyaran (desen) kompleksi 187-188 "
306
beyindeki katekolamin düzeyi üzerindeki etkisi 205—208
ve yatkınlık 185 - 189
sinapsların yapısı 77, 240" 241, 242
İspinoz (Fringilla coelebs) 37
Baskı 39-41
biyokimyasal çalışmalar 48-60, 63-80, 122
yaş değeri 53, 54, 56
ve ilişkisel öğrenme 126-147
yatkınlık 175-197,
306-308
morfolojik sonuçlar 227, 231-255, 302
radyootografik çalışmalar 87-99, 122
hassas dönem 40, 301-305
elektrofizyolojik çalışmalar 228-230, 239, 240-242
Yapay damgalama uyaranları 52, 54, 67, 90, 140, 174, 199-200, 231
ve norepinefrin içeriği
beyinde 206, 212
takviye olarak 130, 133, 138, 139, 141
stimülasyon süresi 71-77
Kalamar (Loligo vulgaris) 27—29"
31
Kalmodulin 246-249
Kalsiyum iyonları 29, 30, 32, 246-247' 248, 249
Kanaryalar (Serinus canarius) 161, Katekolaminler 231, 324, 325
yaşın beynin farklı bölgelerindeki düzey üzerindeki etkisi 205-208
Quintofrontal yol 270, 323 Nucifraga caryocataces
telensefalon 313, 314-318
Korteks 315, 316
Kediler 241, 264, 297
nöronların plastisitesi 204, 205, 213, 302-304
bağımlılık 14, 16, 17, 25, 36
Tavşanlar 17-20, 36, 297
Sıçanlar 96, 110, 203, 208, 220, 254, 295
Beynin yan bölgesi (bölgesi) 97
hasar 109, yazılım, 114, 120, 145, 275
döngü 320, 321
Ligandlar 246
Radyoaktif lizin 50-58, 61, 69-70
Luteinizan Hormon 220
Rhesus maymunları 265, 299-301, 307
Medial septal çekirdek 249 Metil antranilat 143-144, 234 Metiyonin enkefalin 278
Anterograd taşıma yöntemi 261
retrograd taşıma 260 - 216
Memeliler 39, 40, 275, 324, 325
kuşlarla karşılaştırma 280-285, 295, 327
Beyin, tavuk yapısı 312-318 Beyincik 272, 312, 313, 314 Monoaminler 202-203, 324-325 Motivasyon 122, 170
Motor nöronlar (motor nöronlar) 24, 25, 171, 251, 252
Motor korteks 329
Multisinaptik yollar 24, 265-266
Fareler 77, 208, 241-242
Dış krank gövdesi 17, 241, 264
Nörotransmiterler 277-278
ve bağımlılık 27, 29, 30-32
Reseptörler 240, 246, 249, 252 Nöromodülatörler 31, 246, 277-279 Nöronlar 13, 25, 279-280, 307
plastisite 204-205, 213, 302-304
bağımlılık 17-23, 33-37
düzenli taburcu 297-298
yapısal değişiklikler 77, 81,
227, 231-255
işlevsel bağlantıların kurulması 221, 254, 306, 307
fizyolojik değişiklikler 228-230
Nöropeptidler 158, 278
Neokorteks 92, 158
Neostriatum 55, 91, 92, 97, 98, 113, 315, 316, 317
PMVP 262, 263, 270, 276, 279 ile iletişim
işitsel yollar 95, 320, 322
somatosensoriyel yollar 269-270, 322
Nöral tüp 312, 314
Alt temporal lob 121, 135, 138
havlama 265, 266, 280
Norepinefrin 203-205, 246, 248
baskı üzerindeki etki 205, 206-214, 220, 222, 245, 248
Maymunlar, tanıma testi 137-138; 144, 294-295
Koku ampulü 270, 271, 313, 314, 323
sistem 270, 271, 323
Koku alma uyaranları 40
Eğitim 14, 38, 48
ve yatkınlık 185, 195, 196, 306-308
65-67, 129, 135, 266, 275, 295, 301 formlarının ayrımına (farklılaşmasına ) göre
Öğrenme eğrisi 71-73
Oval çekirdek 320, 321
Yuvarlak çekirdek 263, 317, 318
Edimsel öğrenme 130-135, 141-143, 146, 181, 193
Yaklaşık reaksiyon ( refleks) 14, 35
Paleostriatum yükseltilmiş 55, 315, 316, 317, 319
katekolamin içeren yollar 325 326
'PMWG 273, 276, 283 ile bağlantılar
norepinefrin içeriği
fonksiyonlar 165, 273
ilkel 55, 315, 316, 317
Bellek, nörofizyolojik yönler 296-301
referans 166, 168
izler 12, 86, 180, 253, 291
teori 290-296
Parahipokampal bölge 274, 283, 316, 317
Parakoku alanı 315, 323, 326
Periectostriatal kemer 263, 272, 319 320
Alan! 95, 268, 269, 320, 321
Pasif kaçınma 143, 144, 145
Ön beyin 118, 119, 277, 313, 314-318
3 H-lizin 57, 61, 69-70'in dahil edilmesi
3 H-urasil 60, 61, 69-70, 73-76, 78-82, 86
radyootografik çalışmalar 91-92, 95
katekolaminlerin içeriği 205-208, 209, 210, 212
optik kiazma 64, 313, 314, 319
Yaban turpu peroksidaz 260-261
PMVG, sağ ve sol arasındaki etkileşimler 161-165
150, 231-255 sinapslarının yapısındaki değişiklik
hücresel organizasyon 279-280
nöroaktif bileşenler 249, 275-278
tek taraflı imha 113-118, 150-165
hasar 101-103, 171, 213, 292
iki taraflı 101-113, 126-135, 142-143, 175-180, 193-196
sağ ve sol, fonksiyonlar 150-159, 165-170
radyo imzası 95, 97, 99, 143
iletişim 95, 120, 260-277
katekolamin içeriği 205, 212 219 222
fonksiyonlar 122-124, 138-147, 171, 221, 227, 283, 296
ve tercih 175-180, 193-197, 306
elektrik stimülasyonu 199 - 202
elektrofizyolojik çalışmalar 228-230, 239-241, 303
Agresif davranış 273
kaçınma 122, 138, 141, 166, 167, 168, 304
Takviyeler 126, 128, 130, 138, 139, 141, 144, 202, 219
sıcak hava 126-128, 144
Yastık 266
Cinsel damgalama 40
Beynin yarım küreleri 312, 313 bölme 64-71, 95
Postsinaptik membran 233, 240-241, 250-254
kalınlaşma 232, 233-241, 245-249
Prednerv 28, 31
Tercih, çift ayrışma 219
üretim 101-108, 114-115,
116, 119
bireysel varyasyonlar 78-82
ve PMVG 101-108, 113-118'e hasar
koruma 108-118, 146, 151
testler 67-69, 97-98, 103-104, 133, 134
duyarlılık 80, 188-189 Yatkınlık 174-189
ve PMVG 175-180, 306
X' 196
ne için? 189-192
davranış analizi 180-189 Presinaptik tomurcuklar 232, 235 Prefrontal korteks 281, 282, 283,
284, 294-295
Yaklaşım 79, 105-107, 228
ve 3 H-urasil 79, 80, 81, 90 alımı
Alışma 14-39, 41, 48, 86, 291, 293, 304, 305
uyaran genellemesi 18—19, 34
hücresel mekanizmalar 23-33
davranış özellikleri 14-15, 37-39
varsayılan sinir ağları 33-37
hassaslaşma 31-33, 38
kendiliğinden düzelmesi 20-21, 31
fizyolojik analiz 15-23 Primatlar 281, 294-296 Düz mavi 261
Kuluçka kuşları 39, 41
beyin 312-329
ilahiler 161, 219, 307
memelilerle karşılaştırma 280-285, 295
Radyootografik çalışmalar 87-99, 261
sapının retiküler oluşumu 32
Reseptörler 240, 246, 249, 253, 254
Pembe yanaklı muhabbet kuşları (Agaropornis roseicollis) 328
RNA, değişim 58-59, 61, 67, 69, 70, 95, 227, 249
Schistocerca gregaria) 21-22, 30
Sensör sistemleri 262-271, 318-323
Septal çekirdek(ler) 283, 315, 316, 317
— iç organ-endokrin fonksiyonlar 274, 275
yanal 274, 275
kolinerjik bağlantılar 277, 278
Sinapslar 13, 77
ve bağımlılık 24-33
değişiklik 48-49, 231-255
işlevsiz 254, 306, 307
Sinaptik tomurcuklar 233, 235
Mavi nokta 202, 203, 271, 272, 324, 325, 326
Protein sentezi 49-58, 60, 227, 249
SMVG 179, 193, 196 dışında tanıma sistemi
Sığırcık (Sturnus vulgaris) 321, 322
Kabuk 325
İşitsel uyarım ve damgalama 40, 41, 90, 95, 102, 211, 269
- bağımlılık yapan 18, 19, 35
Baykuşlar 263, 303, 319, 320, 328-329
Omurilik, refleksler 13, 15, 24-25, 29, 254
Orta beyin 55, 56, 57, 60, 313, 314
dorso-lateral çekirdek 320, 321
lastik 325, 327
Fleksiyon refleksi (refleks uzuv fleksiyonu) 24, 29
Beyin sapı 203, 271, 312, 313, 314, 320, 321, 323-327
Stres 63, 64, 70, 274
Çizgili kabuk 266
Alt ders 283
Talamofugal yol 318, 319, 320 Talamus 202, 264, 281, 294, 295, 314, 318
- ana görsel çekirdek 263, 319
Tektofugal yol 263, 318, 319 Peck testi 106, 112, 113 HO tanıma, 111, 134-135, 137 138
Testosteron 214-222, 254, 306, 307 İnhibitör postsinaptik potansiyel (IPSP) 27.
Tanıma 134-135, 294-295
— ve ilişkisel öğrenme 135—
138, 144
PMVG 134, 144, 146, 150, 153-159, 168, 193-196, 227, 295
kasa X' 155-156, 157, 168, 196, 296
hipokampusun rolü 144, 284
Urasil, imza 87-95
içerme 58-60, 61, 227
bölünmüş beyine 64-71
ve görsel uyarım 73-78, 122, 250
Koşullu refleksler 39, 136, 137, 250
Ördekler 192, 323, 328
Ördek yavrusu 53-54, 130, 139, 175, 185, 269
Fornix, Hasar 144, 293-294
X' ve doldurulmuş tavuk tercihi 196
Kolesistokinin 278
Bilgi depolama 48-49, 165-170, 179
ve morfolojik değişiklikler 997 9?1 94P
- PMVG 123, 143, 295, 150-159, 227, 165-170'teki izler
Storage X ve Sinir Modeli 161-165
Kronik alkolizm, amnezi 136
tavuğu (Numidia meleagris) 95, 97-98, 269, 272, 321
Döngüsel AMP 246
Cingular kabuğu 281, 283, 284, 294, 295
Tavuk, testosteronun damgalama üzerindeki etkisi 214-219, 220-222
baskı 39-41, 303-305. Ayrıca bkz .
katekolamin içeriği 205-214, 219-220, 221-222
beyindeki nörotransmiterler 278, 327
Dikenler dendritik 77, 233, 235, 241-243, 247
Spike aparatı 247
İnsan, hafıza kaybı 135-137, 283, 290-292, 296
— bağımlılık yapan 35, 293
serebral korteksin tek taraflı lezyonları 121 , 161
beyin sapı fonksiyonları 323-325 Kranial sinirler, beşinci çift
270, 323
Kaşıma refleksi 15, 23-24
Doldurulmuş tavuk ve saklama sistemi X' 196
Ektostriatum 200-201, 315, 316, 317
- görsel yollar 202, 263, 318 -
319
— radyo imzası 91, 92 Elektrik stimülasyonu 200-201, 242, 260
Elektron mikroskobik incelemeler 231, 232, 233-240
Endokrin fonksiyonlar, düzenleme 274
Nükleer sarı 261
Çapraz kirişin çekirdeği (n. a & cumbens) 325-327
Japon bıldırcını (Cotirnix coturnix japanica) 241
İÇİNDEKİLER
Çevirmenin önsözü 5
Yazarın Rusça baskıya önsözü $
Önsöz 10
Bölüm 1. LA'NIN NÖRONAL BAZLARININ ANALİZİNE YAKLAŞIMLAR
MİYATI 12
Bellek ve alışkanlık 14
Baskının özellikleri 30
Devam 41
edebiyat 42
Bölüm 2. İZLEMENİN BİYOKİMYASAL ETKİLERİ ... 48
Protein metabolizması 50
RNA değişimi 58
Yorumlamada bazı zorluklar 60
Devam 62
Edebiyat 62
Bölüm 3. BİYOKİMYASAL DEĞİŞİKLİKLER ÖĞRENMEDE MİDİR? 63
Bölünmüş beyin 64
Duyusal uyarımın bazı etkilerinin kontrol edilmesi ... 71
Biyokimyasal ve davranışsal parametrelerde bireysel farklılıklar 78
Yorum 82
Devam 82
Edebiyat 83
Bölüm 4. BEYİNDE YERLEŞTİRME 88
Radyoaktif urasil kullanarak araştırma yapın . . 88
Radyoaktif 2-deoksiglukoz 96 kullanarak çalışma
Bazı çözülmemiş sorunlar 98
Devam 99
edebiyat 100
Bölüm 5
BELLEK PARÇALARI 101
PMVG hasarı ve tercih gelişimi . . 101
PMVG Hasarı ve Tercihin Korunması .... 108
PMIG'nin tek taraflı yıkımının oluşum üzerindeki etkisi
ve bellek izlerinin saklanması 118
Beyin Yaralanmalarını Kullanan Diğer Araştırmalar. 118
Beyin İmha Verilerinin Değerlendirilmesi 121
Devam 124
Edebiyat 124
Bölüm 6. BASKI VE İLİŞKİLİ ÖĞRENME 126
Sıcak hava ile takviye edildiğinde reaksiyon gelişimi . 126
Görme yeteneği ile güçlendirildiğinde tepkiler geliştirmek
nesne 130
Tanıma ve İlişkisel Öğrenme. . 135
Baskının belirli bir bağımlılığı var mı?
PMVG mi? 138
Devam 147
Edebiyat 147
Bölüm 7. KAÇ İZ DEPOLAMA SİSTEMİ? BEYİN ASİMETRİSİ VE İZLEME 150
Art arda hasar veren deneyler 150
Hemisferik işlev değişikliği 159
Kaç tane hafıza deposu var ve bunlar ne için? 165
Beyin hasarı, motivasyon ve iletişim yolları
Timul ile - reaksiyon 170
Devam 171
Edebiyat 171
Bölüm 8. ETKİLEŞİM VE TERCİH 174
Beyin Hasarı ve Tercihi 175
Davranışsal Tercih Analizi 180
Neye yatkınlık? 189
Doldurulmuş tavuğun tercih edilmesinde PMVG'nin rolü nedir? . . 193
Depolama sistemi X' ve doldurulmuş tavuk 196
Devam 196
Edebiyat 197
Bölüm 9 BELLEĞİN FİZYOLOJİK SINIRLARI VE KİM YAZDIRMA 199
Beyin stimülasyonunun etkileri 199
Yaş ve Görme Deneyiminin Katekol Düzeylerine Etkisi
tavuk beyninin farklı bölümlerinde yeni 205
Norepinefrin ve damgalama 208
Testosteron ve damgalama 214
Çift ayrışma 219
Yorumlar ve akıl yürütme . . 219
Devam 222
Edebiyat 223
10. Bölüm
ÖĞRENME SÜRECİYLE 227
Öğrenmenin Fizyolojik Sonuçları 228
Öğrenmenin Morfolojik Sonuçları 231
Yorumlar ve akıl yürütme 240
Devam 255
Edebiyat 255
Bölüm 11
İletişim SMVG'si
PMVG'nin biyokimyası ve hücresel öğeleri
Yorumlar ve akıl yürütme: kuşlarda ve memelilerde sinir yolları
Özet
Edebiyat
Bölüm 12. SENTEZ YOLU ÜZERİNDE . .
Hafıza ....
Baskı ....
Devam ....
edebiyat _ . ....
Başvuru. KUŞLARIN BEYNİ YAPISININ , ÖZELLİKLE TAVUKLARIN İSİMLERİNE İLİŞKİN BİR ÇERÇEVESİ
S.1. Tavuk beyninin genel tanımı
S.2. Telensefalonun iç yapısı
PZ Sensör sistemleri
S.4. Beyin sapının orta kısmının projeksiyonları
S.5. Serebral hemisferlerin inen yolları
Pb Özet
Edebiyat
konu dizini
Not: Bazen Büyük Dosyaları tarayıcı açmayabilir...İndirerek okumaya Çalışınız.
Yorumlar
Yorum Gönder